Efecto
del Pectimorf®
en el índice estomático de plantas de frijol (Phaseolus
vulgaris L.)
Pectimorf®
effect on bean (Phaseolus vulgaris L.) stomatal index
Dra.C. Idioleidys Álvarez Bello, Dra.C. Inés M. Reynaldo Escobar
Instituto Nacional
de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las
Lajas, Mayabeque, Cuba, CP 32 700.
RESUMEN
Pectimorf®
es un producto
bioactivo que potencia el crecimiento y desarrollo de las plantas en condiciones
de sequía. No obstante, se desconoce su mecanismo de acción en
la planta, por lo que la presente investigación tuvo como objetivo determinar
su efecto en la densidad de los estomas en la superficie foliar de plantas de
frijol (Phaseolus vulgaris L.). Para ello se empleó el cultivar
cubano Cuba C 25-9, el cual se asperjó con 10 mg L-1 de Pectimorf®.
Para el estudio anatómico la epidermis foliar fue observada al microscopio
óptico de luz (Motic) y fotografiadas con una cámara acoplada.
Las variables estomáticas fueron medidas mediante el programa morfométrico
ImageJ y procesadas con el paquete estadístico Stargrafic 5.0. Se demostró
que Pectimorf®
provoca un incremento en el índice estomático (IE) del cultivar
estudiado. La estructura de las células epidérmicas permaneció
inalterable en ambas superficies foliares; sin embargo el tamaño de las
células estomáticas se modificó con la presencia del producto.
En la superficie abaxial las células oclusivas se mostraron más
estrechas y cortas, lo que evidenció que las hojas de mayor índice
estomático presentaron estomas más pequeños. Los resultados
permiten aseverar que Pectimorf®
provoca modificaciones en los patrones de distribución y morfogénesis
estomática, lo que pudiera favorece el crecimiento adecuado de las plantas
de frijol en ambientes desfavorables.
Palabras
clave: índice estomático, oligogalacturónidos,
frijol, Phaseolus.
ABSTRACT
Plant development
and growth is potentiate by Pectimorf under drought conditions, but it action
mechanism is unknown. Due to, to determine the effect of Pectimorf®
on stomatal distribution in bean (Phaseolus vulgaris L.) leaf surface
was the aim of this research. C 25-9 Cuban cultivar was employed and sprayed
with Pectimorf®
to rate of 10 mg L-1.To anatomical study, foliar epidermis was observed
to optical microscope (Motic) light and photographed with a camera attached.
Stomata variables were measured by morphometric ImageJ program and processed
with the statistical package Stargrafic 5.0. Stomata index increase was observed
in Pectimorf®
treated plant and this result could be to favour plant water status. The structure
of epidermal cells remained unchanged on both leaf surfaces, however, some changes
in size of stomatal cells were observed in presence of Pectimorf®.
More narrow and short occlusive cells on abaxial surface were exhibited on treated
plants, so leaves with the most stomatal index were characterized by smallest
stomata. The results obtained allowed to know that Pectimorf®
causes changes in the distribution patterns and stomatal morphogenesis, which
could be to favors successfully growing of bean plant in unfavorable environments.
Key words:
stomatal index, oligogalacturonides, bean, Phaseolus.
INTRODUCCIÓN
La superficie
epidérmica foliar presenta un gran número de poros microscópicos
llamados estomas. La apertura de dichos poros se controla a través de
los cambios en el tamaño y la forma de dos células especializadas,
llamadas células oclusivas, que flanquean la apertura estomática
y poseen una estructura característica que les permite regular la apertura
del poro estomático (1). Estas estructuras se encuentran en todas las
partes aéreas de la planta, pero son más abundantes en las hojas,
dado que la epidermis y la cutícula de los órganos aéreos
forman una capa continua y los estomas son las discontinuidades por donde la
planta realiza la mayor parte del intercambio de O2, CO2, vapor de agua y otros
gases (1, 2).
La forma de los estomas es distintiva de los diferentes grupos de plantas, siendo
muy conocida la diferencia entre mono y dicotiledóneas (1). Los patrones
de desarrollo y estructura estomática han sido abordados en diversos
estudios (3, 4, 5), así como la respuesta de los estomas en cuanto a
tamaño, apertura, y densidad en función de la concentración
de CO2 (6, 7), intensidad de la luz (6, 8) régimen hídrico (9,
10) y variación de la temperatura (11).
Según la teoría más reciente que explica el desarrollo
y los patrones de distribución estomáticos, los mismos pueden
formarse a partir de células que poseen la información genética
necesaria, llamadas meristemoides satelitales y que se encuentran en la epidermis
de la hoja (12). De manera que este carácter es susceptible a cambiar
en ciertas etapas de crecimiento de la planta o en respuesta a diversos estímulos
ambientales (13, 14, 15, 16, 17, 18, 19, 20) que sean capaces de desencadenar
divisiones sucesivas de estas células en potencia.
Razones por las cuales, el objetivo del presente estudio fue evaluar el efecto
de Pectimorf®
en la distribución de los estomas en la epidermis foliar de plantas de
frijol (Phaseolus vulgaris L.).
MATERIALES
Y MÉTODOS
La investigación
se realizó en el Departamento de Fisiología y Bioquímica
Vegetal, del Instituto Nacional de Ciencias Agrícolas, empleándose
el cultivar cubano de frijol (Phaseolus vulgaris L.), Cuba C 25-9.
Para el desarrollo del experimento se sembraron un total de 30 plantas en macetas
plásticas de 5 L, en un suelo Ferralítico Rojo compactado (21).
Con el propósito de garantizar una adecuada fijación biológica
del nitrógeno en todas las plantas, se aplicó Azofert a las semillas
en el momento de la siembra, a razón de 200 mL por cada 50 kg de semillas
(22). Para el estudio del efecto de la mezcla de oligogalacturónidos,
15 plantas fueron asperjadas con una solución de Pectimorf®
a razón de 10 mg L-1 al iniciarse la segunda etapa de la fase
vegetativa de cultivo, específicamente, en el momento en el que el 50
% de las plantas tenían desplegada su primera hoja trifoliada.
El muestreo foliar se realizó en las plantas controles y asperjadas,
cuando comenzó el llenado de las vainas y para el mismo se seleccionó
la tercera hoja trifoliada de cinco plantas por tratamiento. Para obtener la
epidermis foliar se siguió el procedimiento de raspado (23). El muestreo
fue siempre en el tercio medio de la hoja, entre las nervaduras y una vez obtenidas
las láminas epidérmicas se procedió a contrastarlas con
azul de toluidina. Posteriormente fueron fotografiadas con una cámara
acoplada al microscopio óptico de luz (Motic), con un objetivo de 40X
(IE) y 100X y un ocular de 10X.
Se realizaron un total de 20 observaciones (400X), correspondientes a cinco
repeticiones por cada tratamiento. La caracterización de los estomas
(1000x) y la determinación del índice estomático (IE) (400x)
se realizaron sobre las microfotografías tomadas al microscopio y con
el empleo del Programa Morfométrico ImageJ.
Para el conteo de estomas y células epidérmicas se consideró
el número de estomas cuando las dos células oclusivas estaban
presentes y las células epidérmicas, cuando tenía más
del 60 % en el área muestreada.
El índice estomático (IE) se calculó utilizando la fórmula
siguiente (24):
IE= [E/ (E+CE)] x 100.
dónde:
E: número de estomas
CE: número de células epidérmicas
La comparación
de medias se realizó por la prueba T de Student, para el 5 % de probabilidad,
empleando el paquete estadístico STATGRAPHICS Versión 5.0 sobre
Windows (25).
RESULTADOS
Y DISCUCIÓN
Se pudo apreciar
una estructura epidérmica similar en plantas de Phaseolus vulgaris
L. asperjadas y en plantas controles, donde los estomas paracíticos,
distribuidos al azar, exhibieron dos células anexas dispuestas paralelamente
con respecto a las oclusivas. Estas características coinciden con la
descripción realizada anteriormente en plantas de frijol común,
donde también las células oclusivas mostraron una forma arriñonada,
como es típico de las plantas dicotiledóneas (Figura
1).
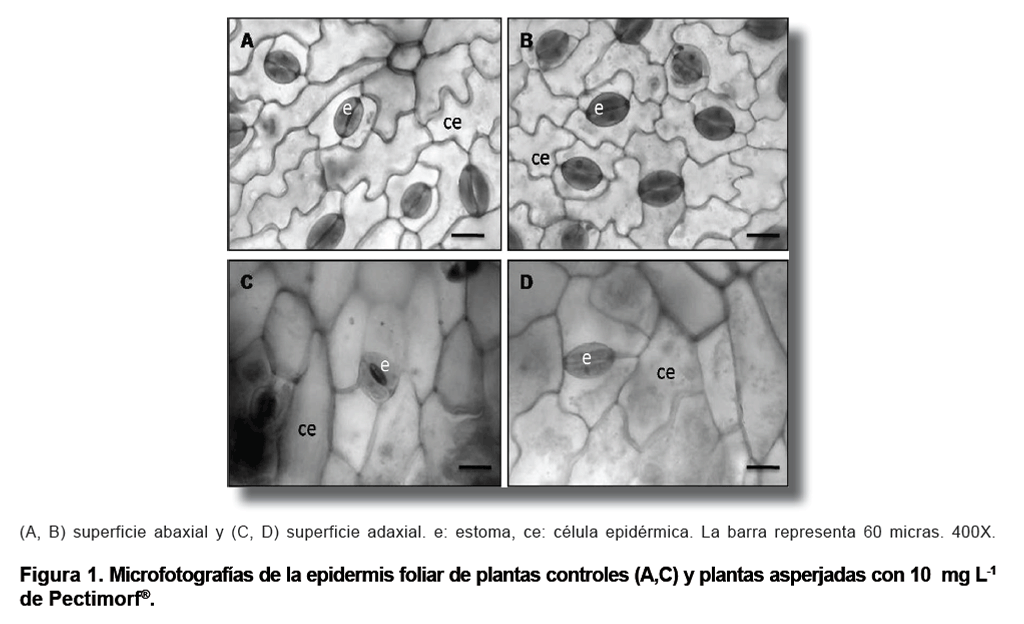
En relación a su localización en hoja, los estomas se pudieron
observar tanto en la superficie abaxial como en la adaxial, por lo que se considera
una especie anfiestomática (1, 26, 27). No obstante, se pudo apreciar
una mayor frecuencia de aparición de estomas en la superficie abaxial
(Figura1). Resultado que responde a un mecanismo que han desarrollado las plantas
para evitar que la luz solar incida directamente sobre los estomas y evitar
así la pérdida de agua por transpiración.
La luz es el estímulo más importante para desencadenar el movimiento
estomático y la respuesta de los estomas está estrechamente relacionada
con la recepción y conversión de la señal luminosa en función
de la actividad metabólica de las células oclusivas (28).
Cuando las plantas se someten a estímulos de luz cortos los estomas pueden
regular la transpiración mediante el mecanismo de apertura y cierre;
sin embargo, cuando la exposición es a largo plazo tiene otras implicaciones
para la planta como la deshidratación (29); de manera que la menor localización
de los estomas en la superficie adaxial provoca un incremento en la resistencia
estomática de esta superficie expuesta directamente a la radiación
solar y así evitar la pérdida de agua.
La Tabla I muestra cómo
en la epidermis abaxial de las hojas de las plantas asperjadas con Pectimorf
® se apreció un incremento significativo del número de estomas
y, por tanto, del índice estomático, que no es más que
un valor diagnóstico que expresa el número de estomas por superficie
foliar en los fragmentos de láminas foliares (30).

Este resultado reviste gran importancia tomando en consideración que
la transpiración y la intensidad de la respiración en las plantas
es directamente proporcional al número y abertura de los estomas (31),
ya que estos indicadores influyen directamente en la regulación del intercambio
gaseoso y la tasa fotosintética en las plantas.
La actividad estomática es una de las estrategias a través de
las cuales las plantas regulan la pérdida de agua y la ganancia de carbono,
ya que los estomas responden rápidamente a los cambios ambientales (32).
Lo anterior permite sugerir que un evento que modifique la densidad estomática
en función de los procesos bioquímicos de la fotosíntesis,
podría ser una vía eficaz para mejorar la producción de
algunos cultivos. En este sentido, los resultados indican que, al menos en estas
condiciones, Pectimorf®
pudiera favorecer la capacidad fotosintética ejerciendo su efecto en
los patrones de desarrollo estomáticos en las plantas de frijol.
Por otro lado, Pectimorf®
funciona como un mensajero químico hormonal que regula los mecanismos
de crecimiento y diferenciación en diferentes cultivos, acelerando el
proceso de crecimiento de las plantas, por lo que este producto pudo haber favorecido
la simbiosis Bradyrhizobium-frijol, como ha sido informado en la aplicación
conjunta de Pectimorf con Azofert y de Azofert con EcoMic en el cultivo de la
soya (33).
Se ha demostrado que los oligogalacturónidos afectan el alargamiento
inducido por auxinas, estimulan la formación de flores y la organogénesis
radical (34, 35), así como el engrosamiento de la pared en células
del periciclo, favoreciendo la división de células que conducen
a la formación de nuevos estomas (36).
El análisis morfométrico de los estomas evidenció cambios
en la morfología de los mismos. En las plantas asperjadas con el producto,
tanto la longitud, como el ancho de las células oclusivas, disminuyeron
significativamente en relación con las no asperjadas (Tabla
II), observándose en las primeras estomas más pequeños.

La forma arriñonada de los estomas parece ser una característica
importante para las plantas de condiciones de sombra húmeda y profunda,
pero su comportamiento dinámico lento podría conducir a un problema
en condiciones de sequía (37). Por tanto se pudiera pensar que los pequeños
estomas observados en las plantas tratadas con Pectimorf®,
pudieran abrir y cerrar más rápidamente, siendo capaces de incrementar
con facilidad la conductancia estomática de la hoja y maximizar la difusión
de CO2 durante condiciones desfavorables para la fotosíntesis
(37), si se tiene en cuenta que, la sección transversal donde tiene lugar
el intercambio gaseoso está determinada, tanto por la abertura estomática
(abertura del poro), como por las características anatómicas de
los estomas (del tamaño y densidad) (38), y que en consecuencia, el efecto
de variables ambientales en la conductancia difusiva del vapor de agua en la
hoja consiste en una dinámica a largo y a corto plazo.
Es posible que Pectimorf®
asperjado en la primera hoja trifoliada, haya favorecido la división
celular en la epidermis de las subsiguientes hojas, tomando en consideración
que los procesos a largo plazo se refieren al período de expansión
de las hojas (semanas) e implican efectos ambientales sobre la capacidad de
los estomas para afinar su apertura, pero también en su tamaño
y densidad (39); mientras que la dinámica a corto plazo (segundos a horas)
es mediada por los ajustes en la apertura del poro que son reversibles (28).
La relación que se demuestra entre la densidad de los estomas y el tamaño
de las células oclusivas, también ha sido observada en otras especies,
de manera que altas densidades estomáticas suelen presentarse con células
oclusivas pequeñas en plantas expuestas a condiciones adversas, aun cuando
cada paso del desarrollo estomático, desde la iniciación hasta
la diferenciación, está altamente organizado y regulado genéticamente
(40).
No obstante, la respuesta del número de estomas y el tamaño de
las células oclusivas a las variables ambientales pueden ser estrictamente
dependientes del tiempo y una de las razones por la cual, los complejos mecanismos
fisiológicos de respuesta estomática no han sido completamente
estudiados (41, 42).
La presente investigación mostró evidencias de que Pectimorf®
puede modificar los patrones de desarrollo estomático, minimizando la
pérdida de agua por transpiración. Por otro lado se encontraron
modificaciones en el tamaño de los estomas, posibilitando una adecuada
relación entre la distribución de estos y el tamaño de
las células oclusivas.
Estos resultados constituyen el primer acercamiento al efecto que puede provocar
Pectimorf®
en la morfogénesis estomática de las plantas y sus potencialidades
para favorecer el desarrollo de estas en condiciones adversas.
BIBLIOGRAFÍA
1. Taiz, L. y
Zeiger, E. Plant Physiology, 4.a ed., edit. Palgrave Macmillan, 1 de
septiembre de 2006, p. 764, ISBN 978-0-87893-856-8.
2. Clément, M.; Leonhardt, N.; Droillard, M.-J.; Reiter, I.; Montillet,
J.-L.; Genty, B.; Laurière, C.; Nussaume, L. y Noël, L.D. ‘‘The
Cytosolic/Nuclear HSC70 and HSP90 Molecular Chaperones Are Important for Stomatal
Closure and Modulate Abscisic Acid-Dependent Physiological Responses in Arabidopsis’’,
Plant Physiology, vol. 156, no. 3, 1 de julio de 2011, pp. 1481-1492,
ISSN 0032-0889, 1532-2548, DOI 10.1104/pp.111.174425, [PMID: 21586649].
3. Bergmann, D.C. y Sack, F.D. ‘‘Stomatal Development’’, Annual Review of
Plant Biology, vol. 58, no. 1, 2007, pp. 163-181, ISSN 1543-5008, DOI 10.1146/annurev.arplant.58.032806.104023,
[PMID: 17201685].
4. Casson, S.A. y Hetherington, A.M. ‘‘Environmental regulation of stomatal
development’’, Current Opinion in Plant Biology, vol. 13, no. 1, febrero
de 2010, pp. 90-95, ISSN 1369-5266, DOI 10.1016/j.pbi.2009.08.005.
5. Ogaya, R.; Llorens, L. y Peñuelas, J. ‘‘Density and length of stomatal
and epidermal cells in «living fossil» trees grown under elevated
CO2 and a polar light regime’’, Acta Oecologica, vol. 37,
no. 4, julio de 2011, pp. 381-385, ISSN 1146-609X, DOI 10.1016/j.actao.2011.04.010.
6. Mott, K.A. ‘‘Opinion: Stomatal responses to light and CO2 depend
on the mesophyll’’, Plant, Cell & Environment, vol. 32, no. 11,
1 de noviembre de 2009, pp. 1479-1486, ISSN 1365-3040, DOI 10.1111/j.1365-3040.2009.02022.x.
7. Zhou, Y.; Jiang, X.; Schaub, M.; Wang, X.; Han, J.; Han, S. y Li, M.-H. ‘‘Ten-year
exposure to elevated CO2 increases stomatal number of Pinus koraiensis
and P. sylvestriformis needles’’, European Journal of Forest Research,
vol. 132, no. 5-6, 18 de agosto de 2013, pp. 899-908, ISSN 1612-4669, 1612-4677,
DOI 10.1007/s10342-013-0728-8.
8. Canová, I.; Durkovic, J.; Hladká, D. y Lukácik, I. ‘‘Changes
in stomatal characteristics and photochemical efficiency during leaf development
in six species of Sorbus’’, Photosynthetica, vol. 50, no. 4, 21 de
septiembre de 2012, pp. 635-640, ISSN 0300-3604, 1573-9058, DOI 10.1007/s11099-012-0063-1.
9. Lake, J.A. y Woodward, F.I. ‘‘Response of stomatal numbers to CO2
and humidity: control by transpiration rate and abscisic acid’’, New Phytologist,
vol. 179, no. 2, 1 de julio de 2008, pp. 397-404, ISSN 1469-8137, DOI 10.1111/j.1469-8137.2008.02485.x.
10. Xu, Z. y Zhou, G. ‘‘Responses of leaf stomatal density to water status and
its relationship with photosynthesis in a grass’’, Journal of Experimental
Botany, vol. 59, no. 12, 1 de septiembre de 2008, pp. 3317-3325, ISSN 0022-0957,
1460-2431, DOI 10.1093/jxb/ern185, [PMID: 18648104].
11. Zhao, X.; Dai, X.; Wang, G.; Shen, Z.; Zhang, H. y Qiu, M. ‘‘Developmental
mechanism and distribution pattern of stomatal clusters in Cinnamomum camphora’’,
Russian Journal of Plant Physiology, vol. 53, no. 3, 1 de mayo de 2006,
pp. 310-315, ISSN 1021-4437, 1608-3407, DOI 10.1134/S1021443706030046.
12. Bergmann, D.C.; Lukowitz, W. y Somerville, C.R. ‘‘Stomatal Development and
Pattern Controlled by a MAPKK Kinase’’, Science, vol. 304, no. 5676,
6 de abril de 2004, pp. 1494-1497, ISSN 0036-8075, 1095-9203, DOI 10.1126/science.1096014,
[PMID: 15178800].
13. Salas, J.A.; Sanabria, M.E. y Pire, R. ‘‘Variación en el índice
y densidad estomática en plantas de tomate (Lycopersicon esculentum
Mill.) sometidas a tratamientos salinos’’, Bioagro, vol. 13, no. 3, 2001, pp.
99–104.
14. Peterson, K.M.; Rychel, A.L. y Torii, K.U. ‘‘Out of the Mouths of Plants:
The Molecular Basis of the Evolution and Diversity of Stomatal Development’’,
The Plant Cell, vol. 22, no. 2, 1 de febrero de 2010, pp. 296-306,
ISSN 1040-4651, 1532-298X, DOI 10.1105/tpc.109.072777, [PMID: 20179138].
15. Pillitteri, L.J. y Torii, K.U. ‘‘Mechanisms of Stomatal Development’’, Annual
Review of Plant Biology, vol. 63, no. 1, 2012, pp. 591-614, ISSN 1543-5008,
1545-2123, DOI 10.1146/annurev-arplant-042811-105451, [PMID: 22404473].
16. Salvatori, E.; Fusaro, L.; Mereu, S.; Bernardini, A.; Puppi, G. y Manes,
F. ‘‘Different O3 response of sensitive and resistant snap bean genotypes (Phaseolus
vulgaris L.): The key role of growth stage, stomatal conductance, and PSI
activity’’, Environmental and Experimental Botany, vol. 87, marzo de
2013, pp. 79-91, ISSN 0098-8472, DOI 10.1016/j.envexpbot.2012.09.008.
17. Vodnik, D.; Hladnik, J.; Vrešak, M. y Eler, K. ‘‘Interspecific variability
of plant stomatal response to step changes of [CO2]’’, Environmental
and Experimental Botany, vol. 88, abril de 2013, (ser. Plant Functioning
in a Changing Global and Polluted Environment), pp. 107-112, ISSN 0098-8472,
DOI 10.1016/j.envexpbot.2011.12.029.
18. Pereira, E.G.; Oliva, M.A.; Rosado-Souza, L.; Mendes, G.C.; Colares, D.S.;
Stopato, C.H. y Almeida, A.M. ‘‘Iron excess affects rice photosynthesis through
stomatal and non-stomatal limitations’’, Plant Science, vol. 201-202,
marzo de 2013, pp. 81-92, ISSN 0168-9452, DOI 10.1016/j.plantsci.2012.12.003.
19. O’Carrigan, A.; Hinde, E.; Lu, N.; Xu, X.-Q.; Duan, H.; Huang, G.; Mak,
M.; Bellotti, B. y Chen, Z.-H. ‘‘Effects of light irradiance on stomatal regulation
and growth of tomato’’, Environmental and Experimental Botany, vol.
98, febrero de 2014, pp. 65-73, ISSN 0098-8472, DOI 10.1016/j.envexpbot.2013.10.007.
20. Yang, X.; Yang, Y.; Ji, C.; Feng, T.; Shi, Y.; Lin, L.; Ma, J. y He, J.-S.
‘‘Large-scale patterns of stomatal traits in Tibetan and Mongolian grassland
species’’, Basic and Applied Ecology, vol. 15, no. 2, marzo de 2014,
pp. 122-132, ISSN 1439-1791, DOI 10.1016/j.baae.2014.01.003.
21. Hernández, A.; Pérez, J.; Bosch, D. y Castro, N. Clasificación
de los suelos de Cuba 2015, edit. Ediciones INCA, Mayabeque, Cuba, 2015,
p. 93, ISBN 978-959-7023-77-7.
22. Gutiérrez Cajiao, A.E.; Nápoles García, M.. y Corbera
Gorotiza, J. Medio de cultivo para Bradyrhizobium japonicum, Cuba,
no. CU 22797 A1, no. solc. C12N 1/20, C12N 1/38, 19 de diciembre de 2002.
23. Castellaro G, G.; Squella N, F.; R, U.; Ullrich R., Tamara.; León
C, F. y Raggi S, A. ‘‘Algunas Técnicas Microhistolóigcas Utilizadas
en la Determinación de la Composición Botánica de Dietas
de Herbívoros’’, Agricultura Técnica, vol. 67, no. 1,
marzo de 2007, pp. 86-93, ISSN 0365-2807, DOI 10.4067/S0365-28072007000100011.
24. Shen, Z.; Wang, G.; Liu, Z.; Zhang, H.; Qiu, M.; Zhao, X. y Gan, Y. ‘‘Network
regulation of calcium signal in stomatal development’’, Acta Pharmacologica
Sinica, vol. 27, no. 7, julio de 2006, pp. 950-958, ISSN 1671-4083, DOI
10.1111/j.1745-7254.2006.00400.x.
25. Statistical Graphics Crop STATGRAPHICS® Plus [en línea], versión
5.1, [Windows], 2001 de 1994, (ser. Profesional), Disponible en: <http://www.statgraphics.com/statgraphics/statgraphics.nsf/pd/pdpricing>.
26. Sánchez, M. y Aguirreola, J. ‘‘Relaciones hídricas’’
[en línea], en: Azcon, J. y Talón, M., Fisiología y bioquímica
vegetal, edit. McGraw-Hill Interamericana Editores, S.A. de C.V., 1993, pp.
49-90, ISBN 978-84-486-0033-4, [Consultado: 15 de julio de 2015], Disponible
en: <https://books.google.es/books/about/Fisiolog%C3%ADa_y_bioqu%C3%ADmica_vegetal.html?hl=es&id=4b12AAAACAAJ>.
27. Bonfim, K.; Faria, J.C.; Nogueira, E.O.P.L.; Mendes, É.A. y Aragão,
F.J.L. ‘‘RNAi-Mediated Resistance to Bean golden mosaic virus in Genetically
Engineered Common Bean (Phaseolus vulgaris)’’, Molecular Plant-Microbe
Interactions, vol. 20, no. 6, 30 de mayo de 2007, pp. 717-726, ISSN 0894-0282,
DOI 10.1094/MPMI-20-6-0717.
28. Chen, C.; Xiao, Y.-G.; Li, X. y Ni, M. ‘‘Light-Regulated Stomatal Aperture
in Arabidopsis’’, Molecular Plant, vol. 5, no. 3, mayo de 2012, pp.
566-572, ISSN 1674-2052, DOI 10.1093/mp/sss039.
29. Hachez, C.; Ohashi-Ito, K.; Dong, J. y Bergmann, D.C. ‘‘Differentiation
of Arabidopsis Guard Cells: Analysis of the Networks Incorporating the Basic
Helix-Loop-Helix Transcription Factor, FAMA’’, Plant Physiology, vol.
155, no. 3, 1 de marzo de 2011, pp. 1458-1472, ISSN 0032-0889, 1532-2548, DOI
10.1104/pp.110.167718, [PMID: 21245191].
30. Ripley, B.S.; Gilbert, M.E.; Ibrahim, D.G. y Osborne, C.P. ‘‘Drought constraints
on C4 photosynthesis: stomatal and metabolic limitations in C3 and C4 subspecies
of Alloteropsis semialata’’, Journal of Experimental Botany, vol. 58,
no. 6, 1 de abril de 2007, pp. 1351-1363, ISSN 0022-0957, 1460-2431, DOI 10.1093/jxb/erl302,
[PMID: 17322550].
31. Meng, F.; Peng, M.; Pang, H. y Huang, F. ‘‘Comparison of photosynthesis
and leaf ultrastructure on two black locust (Robinia pseudoacacia L.)’’, Biochemical
Systematics and Ecology, vol. 55, agosto de 2014, pp. 170-175, ISSN 0305-1978,
DOI 10.1016/j.bse.2014.03.025.
32. Tanaka, Y.; Sugano, S.S.; Shimada, T. y Hara-Nishimura, I. ‘‘Enhancement
of leaf photosynthetic capacity through increased stomatal density in Arabidopsis’’,
New Phytologist, vol. 198, no. 3, 1 de mayo de 2013, pp. 757-764, ISSN
1469-8137, DOI 10.1111/nph.12186.
33. Ayala-Boza, P.J. ‘‘Efecto de biofertilizantes y Pectimorf en la producción
de soya (Glycine max L.) en condiciones de secano’’, Revista Granma Ciencia,
vol. 17, no. 2, 2013, ISSN 1027-795X, [Consultado: 4 de abril de 2015], Disponible
en: <http://www.grciencia.granma.inf.cu/vol%2017/2/2013_17_n2.a5.pdf>.
34. Bellincampi, D.; Dipierro, N.; Salvi, G.; Cervone, F. y Lorenzo, G.D. ‘‘Extracellular
H2O2 Induced by Oligogalacturonides Is Not Involved in the Inhibition of the
Auxin-Regulated rolB Gene Expression in Tobacco Leaf Explants’’, Plant Physiology,
vol. 122, no. 4, 2000, pp. 1379-1386, ISSN 0032-0889, 1532-2548, DOI http://dx.doi.org/10.1104/pp.122.4.1379.
35. Vallarino, J.G. y Osorio, S. ‘‘Signaling role of oligogalacturonides derived
during cell wall degradation’’, Plant Signaling & Behavior, vol.
7, no. 11, 1 de noviembre de 2012, pp. 1447-1449, ISSN null, DOI 10.4161/psb.21779.
36. Yin, X.; Wang, J.; Duan, Z.; Wen, J. y Wang, H. ‘‘Study on the stomatal
density and daily change rule of the wheat’’, Chinese Agricultural Science
Bulletin, vol. 22, 2006, pp. 237–242, ISSN 1000-6850.
37. Yan, F.; Sun, Y.; Song, F. y Liu, F. ‘‘Differential responses of stomatal
morphology to partial root-zone drying and deficit irrigation in potato leaves
under varied nitrogen rates’’, Scientia Horticulturae, vol. 145, 20
de septiembre de 2012, pp. 76-83, ISSN 0304-4238, DOI 10.1016/j.scienta.2012.07.026.
38. Taylor, S.H.; Franks, P.J.; Hulme, S.P.; Spriggs, E.; Christin, P.A.; Edwards,
E.J.; Woodward, F.I. y Osborne, C.P. ‘‘Photosynthetic pathway and ecological
adaptation explain stomatal trait diversity amongst grasses’’, New Phytologist,
vol. 193, no. 2, 1 de enero de 2012, pp. 387-396, ISSN 1469-8137, DOI 10.1111/j.1469-8137.2011.03935.x.
39. Savvides, A.; Fanourakis, D. y van Ieperen, W. ‘‘Co-ordination of hydraulic
and stomatal conductances across light qualities in cucumber leaves’’, Journal
of Experimental Botany, vol. 63, no. 3, 2 de enero de 2012, pp. 1135-1143,
ISSN 0022-0957, 1460-2431, DOI 10.1093/jxb/err348, [PMID: 22121201].
40. Fanourakis, D.; Carvalho, S.M.P.; Almeida, D.P.F.; van Kooten, O.; van Doorn,
W.G. y Heuvelink, E. ‘‘Postharvest water relations in cut rose cultivars with
contrasting sensitivity to high relative air humidity during growth’’, Postharvest
Biology and Technology, vol. 64, no. 1, febrero de 2012, pp. 64-73, ISSN
0925-5214, DOI 10.1016/j.postharvbio.2011.09.016.
41. Lee, J.S.; Kuroha, T.; Hnilova, M.; Khatayevich, D.; Kanaoka, M.M.; McAbee,
J.M.; Sarikaya, M.; Tamerler, C. y Torii, K.U. ‘‘Direct interaction of ligand-receptor
pairs specifying stomatal patterning’’, Genes & Development, vol.
26, no. 2, 15 de enero de 2012, pp. 126-136, ISSN 1549-5477, DOI 10.1101/gad.179895.111,
[PMID: 22241782PMCID: PMC3273837].
42. Franks, P.J.; W. Doheny-Adams, T.; Britton-Harper, Z.J. y Gray, J.E. ‘‘Increasing
water-use efficiency directly through genetic manipulation of stomatal density’’,
New Phytologist, vol. 207, no. 1, 1 de julio de 2015, pp. 188-195,
ISSN 1469-8137, DOI 10.1111/nph.13347.
Recibido: 18 de
septiembre de 2014
Aceptado: 7 de enero de 2015
Dra.C. Idioleidys
Álvarez Bello, Instituto Nacional de Ciencias Agrícolas (INCA),
gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700. Email:
ialvarez@inca.edu.cu