Efecto
de extractos acuosos de Helianthus annuus Lin. sobre el crecimiento
de Solanum lycopersicum Lin.
Effect
of aqueous extracts of Helianthus annuus Lin. on Solanum lycopersicum
Lin. growth
Yaniuska
González Perigó,I Oriela Pino Pérez,II
Ángel Leyva Galán,III Zaida I. Antoniolli,IV
Roberto A. Arévalo,V
Yanara Gómez Matos,I
María I. Pavón RosalesIII
IUniversidad
de Guantánamo, Facultad Agroforestal de Montaña.
IICentro Nacional de Sanidad Agropecuaria (CENSA).
IIIInstituto Nacional Ciencias Agrícolas (INCA), gaveta postal
1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
IVUniversidade Federal de Santa Maria. Rio grande do sul. Brasil.
VSecretaria de Agricultura de Estado de São Paulo. Piracicaba.
Brasil.
RESUMEN
Con el objetivo
de determinar el efecto de extractos acuosos de Helianthus annuus Lin.
sobre el crecimiento de Solanum lycopersicum Lin, se tomaron muestras
de raíz y hojas de plantas recolectadas de un sistema de policultivo
ubicado en un agroecosistema montañoso. Los extractos acuosos de girasol
se obtuvieron a partir de raíz y hojas de plantas recolectadas durante
dos años. Se evaluaron los efectos de estos extractos sobre la germinación,
longitud de la radícula y del hipocótilo de semillas de tomate
certificadas, para ello se ejecutaron nueve tratamientos con cuatro repeticiones
dispuestos en placas Petri. Los extractos de raíz de girasol a los 15,
30 y 45 días y de hojas a los 75 y 90 días de desarrollo inhibieron
la germinación de las semillas de tomate y el crecimiento de la radícula
y el hipocótilo. Los extractos de raíz de girasol con 75 y 90
días, estimularon el crecimiento de la radícula y del hipocótilo
del tomate. Estos resultados demuestran que el girasol produce sustancias químicas
que inhiben el crecimiento del tomate y no se recomienda asociar estos dos cultivos
de forma simultánea.
Palabras
clave: girasol, tomate, radícula, hipocótilo, inhibición.
ABSTRACT
The study was
developed in order to determine the effect of aqueous extracts of Helianthus
annuus Lin on Solanum lycopersicum Lin growth. Roots and leaves
samples of sunflower plants were collected from a policulture system located
in a mountainous agroecosystem and their effects. These effects were evaluated
on certified tomato seeds about germination, radicle and hypocotyl length. Nine
treatments with four replicates were established in Petri plates. The extracts
of sunflower from roots, and from leaves, at 15, 30 and 45 and at 75 and 90
days of development, respectively, inhibited tomato seed germination. Sunflower
root extracts at 75 and 90 days of development stimulated tomato radical and
hypocotyl growth. These results showed that sunflower plant produces chemical
substances inhibiting tomato growth specifically the radicle and the hypocotyl
in the earlier 45 days of growth after germination, and it is not recommended
the association of these two crops simultaneously.
Key words:
sunflower, tomato, radicle, hypocotyl, inhibition.
INTRODUCCIÓN
El cultivo del
girasol (Helianthus annuus L.), perteneciente a la familia Asteraceae,
tiene su origen en América del Norte; en la actualidad se expande a todos
los continentes y cubre un área de aproximadamente 18 millones de hectáreas
en virtud de la calidad de su aceite. Este cultivo es considerado entre los
cuatro de mayor producción de aceite vegetal comestible y su aceite ocupa
el segundo lugar en cuanto al consumo mundial de este tipo de producto (1).
El girasol también posee potencialidades en la obtención de forraje
para la alimentación del ganado vacuno lechero en período poco
lluviosoA. La intensificación de su cultivo se relaciona con su utilización
como materia prima para la producción de biocombustible, en Brasil es
impulsado por la creación del “Programa Nacional de Biodiesel”, que trae
como meta la inserción de la agricultura familiar en la producción
de agroenergíaB (2).
Bajo las condiciones de los agroecosistemas montañosos en Cuba se promueven
los sistemas policulturales y dentro de ellos la introducción de la siembra
del girasol. La presencia de esta especie en el agroecosistema contribuye a
la alimentación humana mediante el aporte de un aceite con valor alimentario
alto y, a la vez, se enriquece la alimentación animal con los residuos
del proceso de extracción. Sin embargo, la introducción del girasol
en asociación con el tomate (Solanum lycopersicum Lin) afectó
el desarrollo de este último cultivo fundamentalmente cuando se asociaron
de manera simultánea (3).
Estudios anteriores demuestran que el girasol puede interferir en el desarrollo
de plantas vecinas (4), aunque los mecanismos responsables por esos efectos
no son bien conocidos, estos probablemente ocurren porque el girasol es fuente
de sesquiterpenoides y otros compuestos con actividad biológica variable
entre genotipos (5). Por tanto se requiere profundizar en el conocimiento de
las relaciones ecológicas de la especie introducida y su potencial como
fuente de compuestos bioactivos, especialmente con actividad alelopática.
Las investigaciones referidas a los efectos alelopáticos entre las plantas
han cobrado mayor importancia después del auge de la agricultura ecológica.
Aunque, los resultados alcanzados en este campo aún son insuficientes
para considerarlos como una solución inmediata a problemas que históricamente
ha presentado la agricultura tropical, por su elevada diversidad de especies
en competencia interespecífica, en cualquier cultivo económicoC.
Los aleloquímicos tienen múltiples funciones dentro del entramado
biótico en el área de distribución original: alelopática,
defensa frente a herbívoros, agentes transportadores de metales o agentes
de simbiosis entre microorganismos del suelo y la planta (6, 7).
Un efecto se puede identificar como alelopático cuando se prueba que
es debido a acciones bioquímicas y no a factores edáficos, climáticos
o de competencia por agua, luz y nutrientes (8). Por lo tanto, el objetivo del
trabajo fue determinar el efecto de extractos de girasol sobre el crecimiento
del tomate, para contribuir a esclarecer si las interacciones químicas
están relacionadas con el efecto negativo marcado y producido en el desarrollo
del tomate por la asociación simultánea de estos cultivos.
MATERIALES
Y MÉTODOS
Recolecta
y procesamiento del material vegetal
Las recolectas se realizaron de septiembre a diciembre del 2009 y de septiembre
a diciembre del 2010 en sistemas de policultivo dentro del agroecosistema montañoso
"Finca la Loma", localidad Limonar de Monte Rous, provincia Guantánamo,
Cuba.
Las raíces de girasol (variedad Caburet) se recolectaron a los 15, 30,
45, 60, 75 y 90 días a partir de la emergencia; las hojas se recolectaron
a los 75 y 90 días. El material vegetal se secó a temperatura
ambiente durante 15 días y se molió en un molino de cuchillas.
Obtención de los extractos
El material vegetal seco y molido se extrajo por maceración con agua
durante cuatro horas (2 g en 100 mL de agua destilada) (9). Cada extracto se
filtró con papel Whatman 1 y se almacenó a una temperatura de
4 °C hasta su evaluación. El contenido de sólidos totales
de 1 mL del extracto se determinó en el analizador de humedad electrónico
marca Santorius MA-45, se realizaron tres repeticiones para cada extracto. Se
calcularon las medias y se analizaron con una prueba de medias de Tukey (p<0,05).
Bioensayo de determinación del efecto sobre el crecimiento
El efecto de los extractos acuosos obtenidos de raíces y hojas recolectados
en diferentes momentos del desarrollo del cultivo de girasol se determinó
en placas Petri (11 cm de diámetro) con papel de filtro Whatman No.1
colocado en el fondo. Se emplearon semillas de tomate (variedad Vita).
Los tratamientos evaluados fueron los siguientes:
H20 destilada (control).
Extracto de raíz de girasol 15 días después de la emergencia.
Extracto de raíz de girasol 30 días después de la emergencia.
Extracto de raíz de girasol 45 días después de la emergencia.
Extracto de raíz de girasol 60 días después de la emergencia.
Extracto de raíz de girasol 75 días después de la emergencia.
Extracto de raíz de girasol 90 días después de la emergencia.
Extracto de hojas de girasol 75 días después de la emergencia.
Extracto de hojas de girasol 90 días después de la emergencia.
Para cada placa Petri, se sumergieron 30 semillas en 2 mL de cada tratamiento
(agua destilada o extracto) durante un minuto. A continuación, se colocaron
sobre papel de filtro Whatman 1 previamente humedecido con 5 mL del extracto
correspondiente (agua en el control). Se utilizaron cuatro réplicas por
tratamiento en un diseño de bloques al azar (9). Las placas Petri se
incubaron a temperatura ambiente. Se determinó el número de semillas
germinadas cada 24 horas y la longitud de la radícula y del hipocótilo
transcurridos siete días. Los datos de longitud de la radícula
y del hipocótilo se presentan como porcentaje de diferencia con el control;
por lo tanto cero representa el control, valores positivos representan estimulación
del parámetro estudiado y valores negativos representan inhibición
(10).
Análisis estadístico
Para las diferencias entre las medias, se llevó a cabo la comparación
de medias con la prueba de Tukey al p<0,05, mediante el paquete estadístico
STATGRAPHICS Plus 5.0 (11).
RESULTADOS
Y DISCUSIÓN
En la Figura
1 se exponen los resultados de la evaluación del efecto de extractos
de raíz y hojas de girasol, recolectadas en diferentes etapas de desarrollo
del cultivo, sobre la germinación de las semillas de tomate. El inicio
de la germinación ocurrió a partir de las 72 horas en el tratamiento
control, 40
y 33 % para las evaluaciones de los años 2009 y 2010 respectivamente,
pero los porcentajes fueron muy bajos para todos los extractos transcurrido
este tiempo.
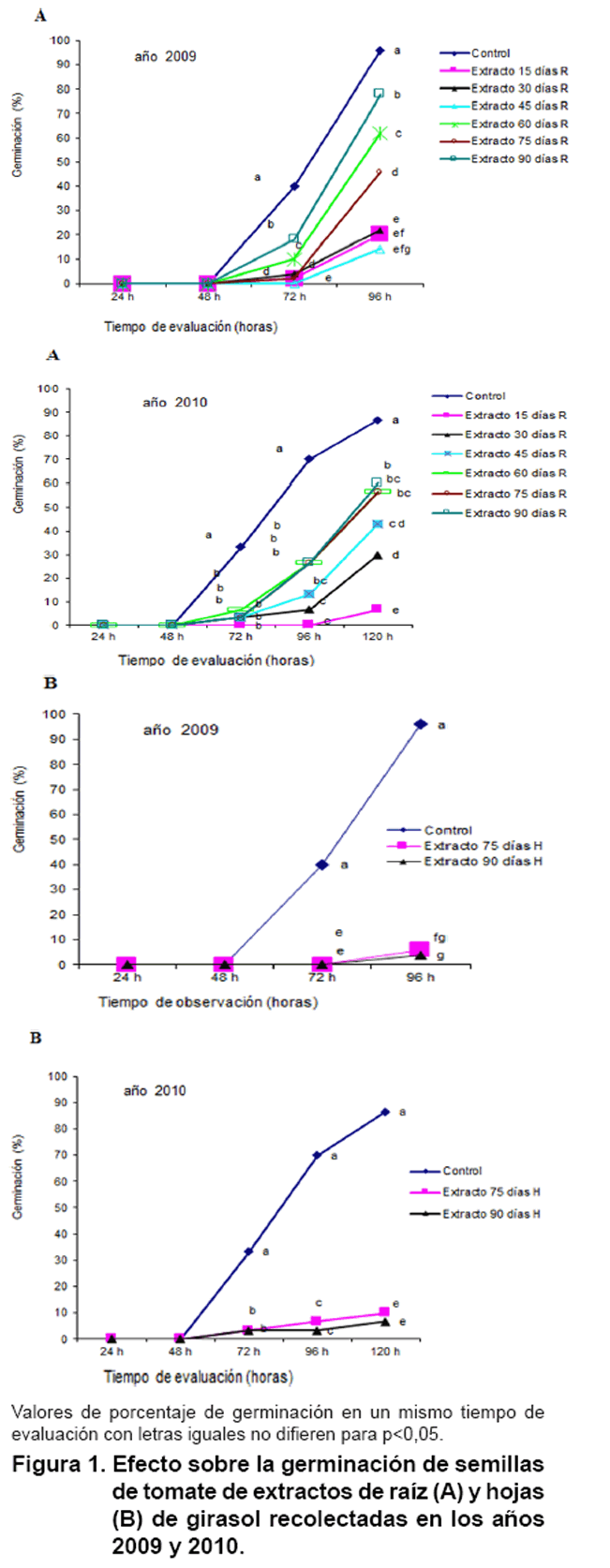
Para el bioensayo del año 2009, la germinación alcanzó
el mayor porcentaje (96 %) en el tratamiento control a las 96 horas, mientras
que el resto de los tratamientos inhibieron significativamente este parámetro.
Para el segundo año, la germinación se monitoreó hasta
las 120 horas cuando el tratamiento control alcanzó el máximo
porcentaje (87 %) y nuevamente el número de semillas germinadas fue menor
en todos los tratamientos que en el control durante todo el tiempo de evaluación
desde el inicio de la germinación.
En ambos años se evidenció que de los extractos de raíz
obtenidos a los 15, 30 y 45 días producen un efecto inhibitorio más
marcado con porcentajes por debajo del 45 %. Cuando se aplican los extractos
de las hojas, los porcentajes son menores del 10 %, inferiores a los obtenidos
en los tratamientos de raíz correspondientes a la misma etapa de desarrollo
fenológico del girasol.
Estos resultados concuerdan con estudios previos que evidenciaron que extractos
acuosos de la parte superior de las plantas, sobre todo hojas, producen una
inhibición mayor para las plántulas que los extractos de raíces
(12).
Diferentes concentraciones de extractos acuosos de hojas de girasol redujeron
la germinación de varias especies como la mostaza (Brassica nigra
(L.) Koch), el rábano (Raphanus sativus Lin.), el trigo (Triticum
vulgare Willd.) y el pepino (Cucumis sativus Lin.) (4). Investigaciones
precedentes también informan sobre la inhibición de la germinación
de otras dicotiledóneas, Lactuca sativa Lin., Lepidium sativum
Lin. y Allium cepa Lin, por compuestos obtenidos de extractos acuosos
de hojas de H. annuus recolectadas un mes antes de la cosecha (coincide
con los 75 días después de la emergencia) (13). También,
diferentes concentraciones de extractos acuosos de hojas de girasol inhibieron
la germinación y el desarrollo de plántulas de trigo (Triticum
aestivum) y mostaza (Sinapis alba), aunque la mostaza fue más
susceptibleD.
La Figura 2 expone los
resultados de la evaluación del efecto sobre el crecimiento de la radícula
de los extractos de raíz y hojas de girasol. Todos los tratamientos del
2009 inhibieron este parámetro independientemente de la parte y la etapa
del desarrollo de la planta. Este comportamiento se repitió con las muestras
del año 2010, excepto para los extractos de raíz de 75 y 90 días
que estimularon el desarrollo radicular.
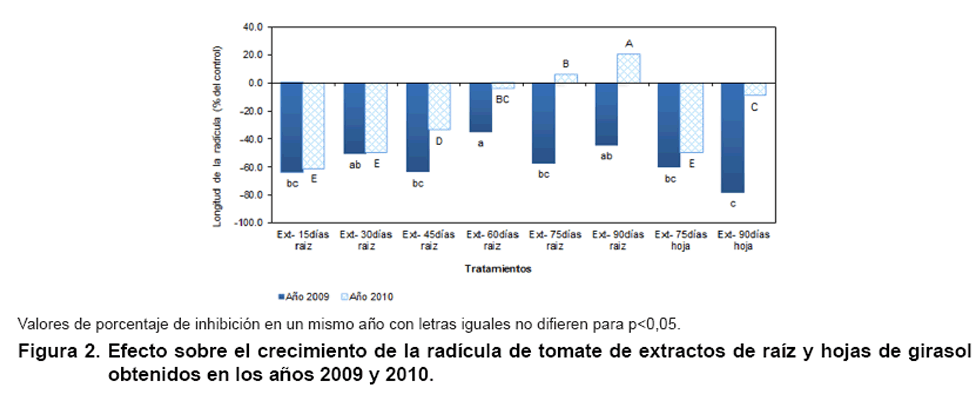
La Figura 3 muestra
los resultados de la determinación del efecto sobre el crecimiento del
hipocótilo de extractos de raíz y hojas de girasol. Las longitudes
del hipocótilo fueron inferiores al control en las variantes a partir
de raíz a los 15, 30, 45 y 60 días, así como las de hojas
a los 75 y 90 días en ambos años evaluados, unido al extracto
de raíz a los 75 días del primer año. Un crecimiento mayor
del hipocótilo, en relación con el control, se produjo en el tratamiento
de extracto de raíz a los 90 días en la evaluación de las
muestras del año 2009; en el año 2010 también, se estimuló
el crecimiento con el extracto de raíz correspondiente a este tiempo
y con el de 75 días.
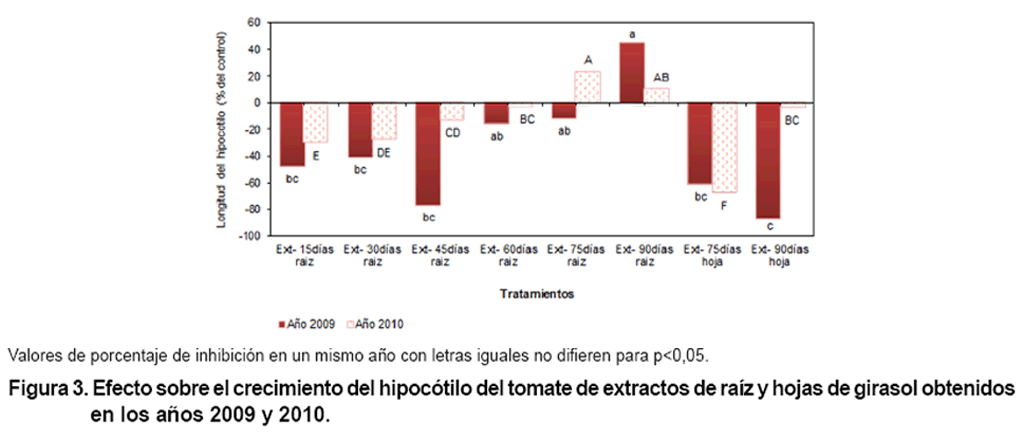
El girasol es una especie conocida por presentar una producción elevada
de metabolitos secundarios; sin embargo, se reconoce la gran variabilidad del
potencial alelopático sobre la germinación y el crecimiento radicular
y la parte aérea entre genotipos de girasol (4, 13). El efecto de sus
metabolitos puede ser estimulador o inhibidor, según el parámetro
evaluado, la especie indicadora utilizada, los diferentes compuestos aislados
a partir de sus extractos y sus concentraciones (13). En este estudio, los efectos
de los tratamientos evaluados sobre los tres parámetros determinados
demuestran la presencia de metabolitos bioactivos en el girasol cultivado (variedad
Caburet) bajo las condiciones del agroecosistema montañoso.
La germinación del tomate se redujo con todos los extractos obtenidos
del girasol durante el crecimiento del girasol y los resultados indican que
las sustancias que se producen al inicio de su desarrollo en la raíz
(hasta los 45 días) y en las hojas en etapas más avanzadas (75
y 90 días) afectan en mayor medida este parámetro, ya sea por
su naturaleza química o por su concentración en las muestras evaluadas.
Además, los compuestos químicos extraídos de las raíces
en etapas tempranas del cultivo desde los 15 a los 60 días y de las hojas
a los 75 y 90 días inhiben no solo la germinación del tomate sino
también el crecimiento y desarrollo de la radícula y del hipocótilo.
Durante los dos años la tendencia de los resultados se mantuvo, en cuanto
al efecto inhibitorio de los extractos obtenidos al inicio del desarrollo radicular
y de las hojas al final del ciclo, lo que podría deberse a que los metabolitos
producidos por el girasol en estas etapas del desarrollo en ambos años
fueron similares.
Por otro lado, el efecto estimulante sobre el crecimiento producido por muestras
de raíz de 75 y 90 días, fundamentalmente en el segundo año
evaluado, indica cambios en los perfiles de metabolitos (cualitativos o cuantitativos)
en las raíces con el desarrollo de la planta. Estos resultados corroboran
la teoría de que los aleloquímicos pueden influir ejerciendo un
efecto inhibitorio o estimulante (14).
Los bioensayos para determinar el efecto de la liberación de compuestos
alelopáticos con frecuencia solo emplean extractos obtenidos, a partir
de la planta donante desarrollada o en proceso de descomposición (10,
15, 16, 17, 18). El diseño experimental utilizado tiene la ventaja de
simular la interferencia girasol-tomate en las diferentes etapas de desarrollo
del girasol. Para lograr la mayor aproximación a la situación
de las plantas en el campo es muy importante representar la interferencia entre
plantas desde los estadios iniciales del desarrollo, momentos en el que el proceso
alelopático puede ser determinante para el establecimiento de las plántulas
como lo demuestra el efecto inhibitorio de los extractos de raíz (de
15 a 60 días de desarrollo) sobre los tres parámetros evaluados.
Estos elementos contribuyen a esclarecer por qué es mayor la afectación
del tomate asociado de manera simultánea con el girasol (3).
Al comparar los resultados de los dos años en cuanto al tipo de efecto
(inhibidor o estimulador) solo se manifiestan diferencias en los extractos de
raíz de 75 días (longitud de radícula e hipocótilo)
y 90 días (longitud de radícula) que pudieran estar asociadas
a variaciones en la biosíntesis de los metabolitos bioactivos, causadas
por cambios en los factores ambientales (clima, suelo y otros) entre un año
y otro. Los efectos alelopáticos de compuestos fenólicos pueden
incrementarse en suelos pobres en nutrientes; por ejemplo, se demostró
que niveles superiores de ácidos cafeoilquínicos en girasol son
causados por deficiencias de nitrógeno, potasio y azufre (19). Los cambios
en el efecto sobre el crecimiento al comparar los resultados de los dos años
estudiados pudieran, por tanto, deberse a variaciones en el contenido de materia
orgánica y de algunos de estos elementos en las áreas de cultivo.
Los mecanismos responsables de los efectos del girasol sobre las diferentes
especies diana estudiadas no están totalmente esclarecidos, pero están
asociados a la biosíntesis de algunos metabolitos entre los que se encuentran
algunas lactonas sesquiterpénicas (5).
Se plantea que la mayoría de esos compuestos ejercen su efecto a través
de un mecanismo de acción común, la alquilación de moléculas
orgánicas.
Los niveles de actividad pueden estar correlacionados con la composición
cuantitativa o cualitativa de los extractos. La variabilidad de la respuesta
de la planta diana puede ser atribuida a diferencias en la concentración
de las sustancias presentes en los extractos de girasol, por ello durante el
segundo año se determinaron los sólidos totales en cada uno de
ellos (Tabla I).

Solo los extractos de raíz a los 15 y a los 90 días poseen valores
de sólidos totales significativamente diferentes; el primero con mayor
concentración inhibe, tanto el crecimiento de la radícula como
del hipocótilo, mientras que el segundo estimula ambos parámetros.
Los valores de sólidos totales de los extractos de raíz no se
correlacionan directamente con el efecto sobre el crecimiento, pues extractos
que inhiben (15, 30 y 45 días) y otros que estimulan (75 y 90 días)
no difieren estadísticamente en su concentración de sólidos
totales. Los extractos de las hojas no difieren entre sí en cuanto a
la concentración de sólidos totales disueltos.
Aunque los valores de los sólidos totales se mantuvieron dentro de un
rango en el que no difieren estadísticamente, las variaciones en el efecto
sobre el crecimiento de los extractos de raíz pueden deberse a que la
composición cualitativa es diferente o a que la cantidad de alguno(s)
de los compuestos bioactivos varía significativamente, aunque el valor
total no lo haga. Los extractos con mayor efecto sobre el tomate pueden tener
una concentración más alta de el(los) compuesto(s) responsable(s)
del efecto biológico. Para que las sustancias alelopáticas ejerzan
su acción sobre las plantas dianas se deben cumplir varias etapas. Los
compuestos deben ser liberados de las plantas donantes en cantidades adecuadas
y alcanzar las concentraciones suficientes para ser absorbidos, traslocados
y actuar sobre algún mecanismo bioquímico relevante para el desarrollo
de la especie diana (19).
La investigación realizada proporciona información muy importante
que contribuye a explicar el marcado efecto negativo de la asociación
simultánea del tomate con el girasol, pues evidencia la presencia de
aleloquímicos que pueden mediar las relaciones ecológicas en el
agroecosistema y confirma la importancia de considerar el impacto ambiental
de la introducción de una especie nueva al sistema.
CONCLUSIONES
El girasol produce
sustancias químicas que inhiben el crecimiento del tomate que pueden
ser responsables del efecto negativo sobre el desarrollo de este cultivo en
asociación.
BIBLIOGRAFÍA
1. Curti, G.L.;
Martin, T.N.; Ferronato, M. de L. y Benin, G. ‘‘Girassol ornamental: caracterização,
pós-colheita e escala de senescência’’, Revista de Ciências
Agrárias, vol. 35, no. 1, junio de 2012, pp. 240-250, ISSN 0871-018X.
2. Pereira, da S.H.; Carvalho, M.G.J. de.; Gomes, N.J.M.; da Silva, B.D.J. y
Karam, D. ‘‘Levantamento das plantas espontâneas na cultura do girassol’’,
Revista Verde de Agroecologia e Desenvolvimento Sustentável,
vol. 5, no. 1, 2010, p. 167, ISSN 1981-8203.
3. González, P.Y.; Leyva Galán, A. y Pino Pérez, O. ‘‘Competencia
por interferencia de Helianthus annuus L., asociado a Solanum lycopersicum
L. bajo condiciones de campo’’, Cultivos Tropicales, vol. 35, no. 4, diciembre
de 2014, pp. 28-35, ISSN 0258-5936.
4. Silva, H.L.; Trezzi, M.M.; Marchese, J.A.; Buzzello, G.; Miotto, J.E.; Patel,
F.; Debastiani, F. y Fiorese, J. ‘‘Determinação de espécie
indicadora e comparação de genótipos de girassol quanto
ao potencial alelopático’’, Planta Daninha, vol. 27, no. 4,
2009, pp. 655-663, ISSN 0100-8358.
5. Macías, F.A.; Marín, D.; Oliveros-Bastidas, A.; Varela, R.M.;
Simonet, A.M.; Carrera, C. y Molinillo, J.M. ‘‘Allelopathy as a new strategy
for sustainable ecosystems development’’, Biological Sciences in Space,
vol. 17, no. 1, 2003, pp. 18–23, ISSN 1349-967X.
6. Oliveros-Bastidas, A. de J.; Macías, F.A.; Fernández, C.C.;
Marín, D. y Molinillo, J.M.G. ‘‘Root exudates and their relevance to
the allelopatic interactions’’, Química Nova, vol. 32, no. 1,
2009, pp. 198-213, ISSN 0100-4042, DOI 10.1590/S0100-40422009000100035.
7. Vázquez, M.L.L. ‘‘Manejo de plagas en la agricultura ecológica’’,
Boletín Fitosanitario, vol. 15, no. 1, 2010, p. 38, ISSN 1816-8604.
8. Lorenzo, P. y González, L. ‘‘Alelopatía: una característica
ecofisiológica que favorece la capacidad invasora de las especies vegetales’’,
Revista Ecosistemas, vol. 19, no. 1, 2010, ISSN 1697-2473, [Consultado:
15 de junio de 2015], Disponible en: <http://revistaecosistemas.net/index.php/ecosistemas/article/view/57>.
9. Caamal-Maldonado, J.A.; Jiménez-Osornio, J.J.; Torres-Barragán,
A. y Anaya, A.L. ‘‘The Use of Allelopathic Legume Cover and Mulch Species for
Weed Control in Cropping Systems’’, Agronomy Journal, vol. 93, no.
1, 2001, p. 27, ISSN 1435-0645, DOI 10.2134/agronj2001.93127x.
10. Macías, F.A.; Lacret, R.; Varela, R.M.; Nogueiras, C. y Molinillo,
J.M.G. ‘‘Isolation and Phytotoxicity of Terpenes from Tectona grandis’’, Journal
of Chemical Ecology, vol. 36, no. 4, 18 de marzo de 2010, pp. 396-404,
ISSN 0098-0331, 1573-1561, DOI 10.1007/s10886-010-9769-3.
11. Statistical Graphics Crop STATGRAPHICS® Plus [en línea], versión
5.1, [Windows], 2000, (ser. Profesional), Disponible en: <http://www.statgraphics.com/statgraphics/statgraphics.nsf/pd/pdpricing>.
12. Chon, S.-U. y Nelson, C.J. ‘‘Allelopathy in Compositae plants. A review’’,
Agronomy for Sustainable Development, vol. 30, no. 2, abril de 2010,
pp. 349-358, ISSN 1774-0746, 1773-0155, DOI 10.1051/agro/2009027.
13. Macías, F.A.; Varela, R.M.; Torres, A. y Molinillo, J.M.G. ‘‘Potential
Allelopathic Activity of Natural Plant Heliannanes: A Proposal of Absolute Configuration
and Nomenclature’’, Journal of Chemical Ecology, vol. 26, no. 9, 1
de septiembre de 2000, pp. 2173-2186, ISSN 0098-0331, 1573-1561, DOI 10.1023/A:1005524601019.
14. Hernández Escobar, I.; Urra Zayas, I.; Díaz Viruliche, L.;
Pérez Petitón, J. y Hernández Cuello, G. ‘‘Labranza mínima
y efecto alelopático en la producción de frijol común en
la Empresa Agropecuaria 19 de Abril de la provincia Mayabeque’’, Revista
Ciencias Técnicas Agropecuarias, vol. 22, no. 3, septiembre de 2013,
pp. 46-48, ISSN 2071-0054.
15. Macías-Rubalcava, M.L.; Hernández-Bautista, B.E.; Oropeza,
F.; Duarte, G.; González, M.C.; Glenn, A.E.; Hanlin, R.T. y Anaya, A.L.
‘‘Allelochemical Effects of Volatile Compounds and Organic Extracts from Muscodor
yucatanensis, a Tropical Endophytic Fungus from Bursera simaruba’’, Journal
of Chemical Ecology, vol. 36, no. 10, 31 de agosto de 2010, pp. 1122-1131,
ISSN 0098-0331, 1573-1561, DOI 10.1007/s10886-010-9848-5.
16. Rodrigues, I.M.C.; Souza Filho, A.P.S.; Ferreira, F.A. y Demuner, A.J. ‘‘Chemical
prospecting of compounds produced by Senna alata with allelopathic activity’’,
Planta Daninha, vol. 28, no. 1, 2010, pp. 1-12, ISSN 0100-8358, DOI
10.1590/S0100-83582010000100001.
17. Casas, L.; Mantell, C.; Rodríguez, M.; Torres, A.; Macías,
F.A. y Ossa, E.J.M.D.L. ‘‘Identification of Major Compounds Extracted by Supercritical
Fluids from Helianthus annuus L. Leaves’’, Solvent Extraction Research
and Development, Japan, vol. 18, 2011, pp. 55-68, ISSN 2188-4765, DOI 10.15261/serdj.18.55.
18. Imatomi, M.; Novaes, P.; Matos, A.P.; Gualtieri, S.C.J.; Molinillo, J.M.G.;
Lacret, R.; Varela, R.M. y Macías, F.A. ‘‘Phytotoxic effect of bioactive
compounds isolated from Myrcia tomentosa (Myrtaceae) leaves’’, Biochemical
Systematics and Ecology, vol. 46, febrero de 2013, pp. 29-35, ISSN 0305-1978,
DOI 10.1016/j.bse.2012.09.005.
19. Vyvyan, J.R. ‘‘Allelochemicals as leads for new herbicides and agrochemicals’’,
Tetrahedron, vol. 58, no. 9, 25 de febrero de 2002, pp. 1631-1646,
ISSN 0040-4020, DOI 10.1016/S0040-4020(02)00052-2.
Recibido: 26 de febrero de 2014
Aceptado: 3 de octubre de 2014
Yaniuska
González Perigó, Universidad de Guantánamo,
Facultad Agroforestal de Montaña.
Email: yaniuska@fam.cug.co.cu