Contenido de Fenoles Totales en frutos de mango 'Super Haden' dañados por antracnosis y tratados en poscosecha
Total phenolic content in 'Super Haden' mango fruits damaged by anthracnose and treated in postharvest
Akira Pérez Márquez,I Yamile Vidal Aguiar,I Tania Mulkay VitónII
IInstituto
Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José
de las Lajas, Mayabeque, Cuba, CP 32700.
IIInstituto de Investigaciones en Fruticultura Tropical, Ave 7ma.
No 3005 e/30 y 32, Playa, La Habana, Cuba.
RESUMEN
El contenido de fenoles totales en el exocarpio de los frutos tiene relación con los mecanismos de defensa propios del fruto frente al ataque por patógenos fúngicos. Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. es causante de la antracnosis en el mango (Mangifera indica L.) y ocasiona elevadas pérdidas en poscosecha. El objetivo de este trabajo fue evaluar el contenido de fenoles totales en frutos de mango ‘Super Haden’ dañados y tratados en poscosecha y evidenciar su relación con los mecanismos de defensa frente a la infección por patógenos. Se tomaron muestras de exocarpios de frutos sanos y con niveles de daños ligeros, moderados y severos por la antracnosis, así como en frutos tratados en poscosecha con agua caliente (53 ºC por 5 min), cera polietilénica (10 % ST) más imazalil (800 mg L-1), cera polietilénica (10 % ST) más imazalil (800 mg L-1) más dos bolsas de Conserver 21 y un testigo sin aplicación. Las muestras se liofilizaron y se determinó el contenido de fenoles totales por el método sugerido por Slinkard y Singleton (1977) con modificaciones. Los frutos afectados por la antracnosis presentaron menores contenidos de fenoles totales en el grado ligero fue de 59,25, en el moderado de 58,63 y en el severo 56,52 mg de ácido gálico g-1 de masa fresca, mostrándose diferencias significativas entre los grados y frutos sanos. Los tratamientos poscosecha incrementaron el contenido de fenoles totales. El agua caliente y la combinación de cera polietilénica con imazalil mostró valores de 37,58 y 37,11 mg de ácido gálico g-1 respectivamente, en comparación con el testigo que fue 33,94 mg de ácido gálico g-1, existiendo una correspondencia con la disminución de la incidencia de la enfermedad.
Palabras clave: antracnosis, exocarpio, fenol, mango.
ABSTRACT
Total phenolic content in the exocarp of fruits is related to their own defense mechanisms against fungal pathogen attack. Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. causes anthracnose in mango (Mangifera indica L.) thereby high postharvest losses. The aim of this study was to evaluate the total phenolic content in damaged and postharvest treated ‘Super Haden’ mango fruits, to make evident its relationship with the defense mechanisms against pathogen infection. Exocarp samples were taken from healthy and anthracnose-damaged (mild, moderate and severe levels) fruits as well as from those postharvest treated with hot water (53 °C for 5 min), polyethylene wax (10 % ST) plus imazalil (800 mg L-1), polyethylene wax (10 % ST) plus imazalil (800 mg L-1) plus two bags of Conserver 21 and a control without application. Samples were lyophilized and the total phenolic content was determined by the method suggested by Slinkard and Singleton (1977) with some modification. Anthracnose-affected fruits showed a lower total phenolic content: 59,25 at the mild degree, 58,63 at the moderate and 56,52 mg of galic acid g-1 fresh weight at the severe level, showing significant differences between damage levels and health fruits. Postharvest treatments increased total phenolic content. Hot water and the combination of polyethylene wax plus imazalil showed values of 37,58 and 37,11 mg of galic acid g-1 fresh weight respectively, compared with the control value of 33,94 mg galic acid g-1 fresh weight, showing a correspondence with a lower disease occurrence.
Key words: anthracnose, exocarp, phenol, mango.
INTRODUCCIÓN
Los compuestos
fenólicos son metabolitos secundarios que se caracterizan por estar formados
por unidades de fenoles-anillos aromáticos unidos a uno o más
grupos hidroxilos. Estas biomoléculas se localizan en todas las partes
de las plantas y su concentración varía a todo lo largo del ciclo
vegetativo; además, participan en diversas funciones, tales como la asimilación
de nutrientes, síntesis proteica, actividad enzimática, fotosíntesis,
formación de compuestos y en la defensa ante factores bióticos
y abióticos (1).
Los mecanismos de defensa activados en las plantas incluyen la producción
rápida de especies reactivas del oxígeno (ROS), alteraciones en
la constitución de la pared celular, la acumulación de metabolitos
secundarios como fitoalexinas, la activación y síntesis de péptidos
y proteínas de defensa, entre otros (2, 3).
Entre los compuestos fenólicos más importantes se distinguen los
ácidos fenólicos, los flavonoides, los taninos y otros compuestos
menos comunes como estilbenos y lignanos (4, 5).
La evolución de los compuestos fenólicos, durante la etapa de
poscosecha o maduración en los frutos, ha sido un tema de interés
para los investigadores, porque pueden estar implicados en los mecanismos de
defensa de los frutos ante la infección de determinados hongos (6, 7).
En ausencia de daños o ataque de patógenos, los mecanismos de
defensa del fruto pueden ser inducidos por elicitores físicos o químicos
(8). Los elicitores físicos incluyen las temperaturas altas y bajas,
las irradiaciones gamma y ultravioletas (9, 10, 11). Entre los químicos
se encuentran productos como el quitosano, benzothiadiazole (BTH), harpin y
1-methylciclopropano (12, 13, 14, 15) y otros como el ácido oxálico
y cloruro de calcio (16).
El hongo Colletotrichum gloeosporioides (Penz.) Penz. & Sacc se
considera muy polimórfico y polífago, por su forma de causar la
infección en los frutos, clasificándose como un hongo de infección
latente, produce enzimas (polygalacturonasa y pectatilyasa) que degradan la
pared celular y ocasiona la enfermedad conocida como la antracnosis en el mango
(Mangifera indica L.) (17, 18, 19).
La incidencia y severidad de los daños por la antracnosis en los frutos
de mango afectan su calidad comercial y provocan pérdidas elevadas durante
la conservación y comercialización (20). Su control en la poscosecha
se efectúa mediante los métodos físico (hidrotérmico)
y químico o la combinación de ambos (21).
En Cuba, el mango es uno de los frutos que más se produce, representando
más del 30 % de la producción total de frutalesA y
existe un gran interés en incrementar su exportación como fruto
fresco, siendo el cultivar 'Super Haden' típico del país y el
más distribuido en todas las regiones productoras, pero las pérdidas
durante la poscosecha son elevadas, debido entre otras causas, a la incidencia
de la antracnosis. Sin embargo, existen escasos estudios relacionados con el
proceso de la infección latente del hongo y el desarrollo de la enfermedad,
durante la maduración de los frutos, así como sus mecanismos de
inducción de resistencia ante la aplicación de los tratamientos
para el control poscosecha de la antracnosis. El objetivo de este trabajo fue
determinar los fenoles totales en frutos de mango 'Super Haden' con diferentes
grados de severidad por la antracnosis y después de tratados en poscosecha.
MATERIALES Y MéTODOS
Los frutos de
mango (Mangifera indica L.) ‘Super Haden’ se recolectaron con madurez
fisiológica de una plantación ubicada en la Unidad Científico-Tecnológica
de Base Alquízar, (UCTB, Alquízar), provincia Artemisa, que se
encuentra en los 82° 32´ N y los 22° 47´ O, a 11 m sobre
el nivel medio del mar, sobre un suelo Ferrasol éutrico con topografía
llana de pendiente 0 (22).
Determinación de fenoles totales en el exocarpio de los frutos
dañados por la antracnosis
Los síntomas por la antracnosis se identificaron según Ploetz
(23) y los daños en el exocarpio de los frutos se determinaron a través
de la escala arbitraria de clasificación según Mulkay et al.
(24). Los resultados se expresaron en el porcentaje de frutos por daños
(Tabla).
Los fenoles totales
en el exocarpio de los frutos clasificados, de acuerdo a la escala arbitraria,
se determinaron por el método colorimétrico de Folin Ciocalteu
(25) con ligeras modificaciones. Se tomaron tres muestras (un fruto por muestra
y por clasificación) y se liofilizaron para su posterior análisis.
Los extractos resultantes se analizaron por espectrofotometría. La absorbancia
se midió con un espectrofotómetro Shimadzu UV-180 a una longitud
de onda de 765 nm y los fenoles totales se calcularon en base a la pendiente
de la proporción lineal de la curva patrón de ácido gálico
y se expresó como mg de ácido gálico g-1 de
masa seca.
Determinación de los fenoles totales en el exocarpio de los frutos tratados
en poscosecha
Los frutos se lavaron con detergente Tropicleaner® 0,01 %, se
enjuagaron y se sometieron a los siguientes tratamientos:
Al final de la frigoconservación se determinaron:
Diseño
experimental
Se empleó un diseño experimental completamente aleatorizado. Para
cada evaluación de los daños por la antracnosis y los tratamientos
se realizaron tres repeticiones (30 frutos x repetición). El procesamiento
estadístico de los resultados se realizó mediante un Anova de
Clasificación Simple. Las medias se compararon por la Prueba de Tukey
(p=0,05).
RESULTADOS Y DISCUSIóN
Determinación
de los fenoles totales en el exocarpio de los frutos dañados por la antracnosis
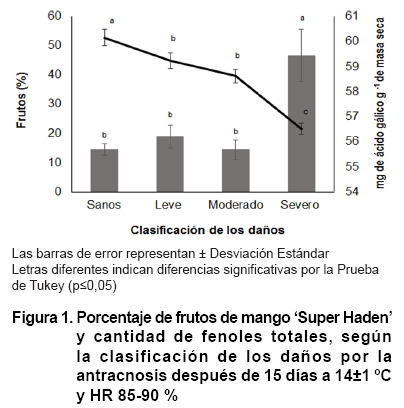
Los frutos de mango se clasificaron como sanos, con daños leves, moderados
y severos por la antracnosis. Sobre el exocarpio se desarrollaron manchas de
forma redonda, hundida, de color pardo oscuro a negro, acompañadas de
una masa de color salmón, como consecuencia de la maduración del
acérvulo del hongo (23). El porcentaje de frutos con daños severos
fue más significativo que el de los daños leves, moderados y sanos.
Al analizar el contenido de fenoles totales en los frutos con daños severos,
hubo menores cantidades con diferencias entre los daños leves y moderados
y estos a su vez con los sanos (Figura 1).
Los resultados
relacionados con el alto porcentaje de frutos de mango con daños severos
por la antracnosis pudiera deberse a la presencia de condiciones favorables
para el desarrollo de la enfermedad, como las temperaturas entre 20 y 30 ºC
por 12 horas y humedad relativa sobre el 95 %, además de la existencia
de abundante fuente de inóculo en la plantación, que favorecen
las infecciones latentes de manera natural del hongo C. gloeosporioides
(27, 28).
La
disminución de los fenoles totales en el exocarpio de los frutos de mango
con daños severos pude ser consecuencia del mayor porcentaje de área
del exocarpio afectada, como resultado de las infecciones latentes del hongo,
además de la presencia de un mayor número de manchas de tamaño
mayor y menor a 1 cm de diámetro, que indican el estado de desarrollo
de la enfermedad, muy diferente a lo observado en los frutos con daños
leves, moderados y sanos, donde las cantidades de fenoles fueron mayores.
Los frutos de mango presentan altos contenidos de compuestos fenólicos
en el exocarpio. Los mayores niveles de estos compuestos se detectan en etapas
juveniles del desarrollo del fruto, produciéndose un importante descenso
durante la maduración.
Los compuestos fenólicos son solamente inducidos cuando los factores
de estrés están presentes. La biosíntesis de las fitoalexinas
suele producirse en zonas localizadas, alrededor del área de entrada
del microorganismo, estando ausente o solo presente en cantidades reducidas
en los tejidos sanos (29).
Algunos autores refieren que los cambios en el contenido de fenoles pudieran
estar asociados al proceso de infección del patógeno y a las características
de los materiales de defensas como la fitoanticipina, recientemente informada
(5).
Cuando la célula de los tejidos está sana e intacta, la enzima
Polifenol Oxidasa (PPO) y sus sustratos, los fenoles se encuentran en compartimientos
separados (cloroplastos y vacuolas, respectivamente). Sin embargo, cuando la
célula se desorganiza, como resultado del proceso infeccioso de la antracnosis
(23), las enzimas y los sustratos se ponen en contacto y sucede la reacción
de oxidación de los compuestos fenólicos, catalizada por la enzima
PPO, causando la formación de quinonas, las cuales son polimerizadas
y forman pigmentos negros, cafés y pardos, afín con los síntomas
de la antracnosis.
Determinación de los fenoles totales en el exocarpio de los frutos
tratados en poscosecha
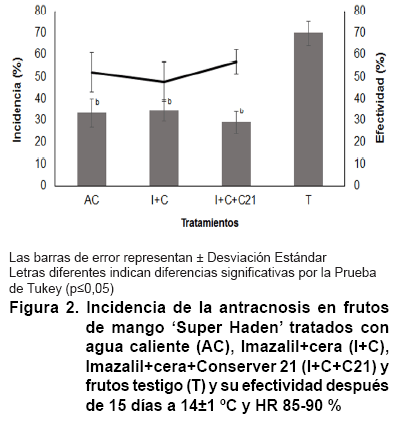
La incidencia de la antracnosis en los frutos de mango, tratados en poscosecha,
disminuyó significativamente; en los testigos la enfermedad afectó
un 66,66 %, por lo que la efectividad de los tratamientos fue superior al 46,87
%, sin diferencias estadísticas (Figura 2).
La efectividad
del agua caliente y el fungicida imazalil para el control de la antracnosis
en frutos de mango, coincide con lo planteado por distintos autores en relación
con el efecto de estos en la calidad y vida de anaquel de diferentes frutosB
(30, 31, 32).
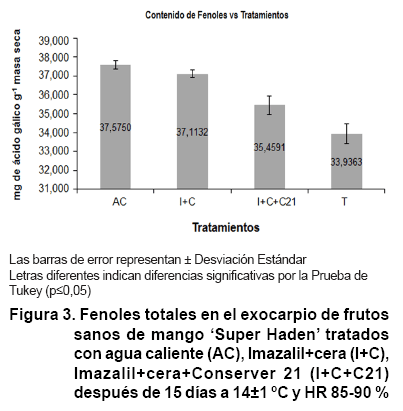
Los tres tratamientos provocaron un significativo incremento del contenido de
fenoles, mientras que en los testigos fue de 33,93 mg de ácido gálico
g-1 de masa seca. El hidrotérmico y la combinación
del fungicida, más la cera, mostraron mayores cantidades de fenoles,
con diferencias estadísticas a la combinación del fungicida, la
cera y el Conserver 21 (Figura 3).
El incremento
de fenoles en el exocarpio de los frutos de mango con agua a 53 ºC x 5
min, son similares a los obtenidos por Talcott et al. (33). Diferentes estudios
señalan los posibles mecanismos del efecto del tratamiento con calor
en la inducción de resistencia a enfermedades y en la calidad poscosecha
de los frutos (34, 35, 36).
Una forma de acción primaria de este tratamiento es la desinfección
del fruto, siendo eliminadas o destruidas las esporas o micelio fúngico
(37).
En frutos de papaya, sumergidos en agua, a 54 ºC x 4 min, hubo una reducción
de los estados de desarrollo de C. gloeosporioides, con una disminución
significativa en la incidencia de la antracnosis, la cera natural de los frutos
se fundió y las grietas y los estomas quedaron cubiertos y sellados,
proporcionando una barrera mecánica contra el ataque del patógeno;
además, este tratamiento indujo la expresión de la proteína
de la polygalacturonasa (38), que puede inhibir la endopolygalacturonasa fúngica,
considerada como un factor importante para la resistencia de las plantas a hongos
fitopatógenos.
También,
la aplicación del tratamiento hidrotérmico a 53 ºC x 3 min
en frutos de melón (Cucumis melo L.), redujo las pudriciones
causadas por diferentes hongos, hubo una mayor actividad de las enzimas fenilalanina
amonoliasa, cinnamata-4-hidroxilasa, coumarata, CoA ligasa, polifenoxidasa y
peroxidasa, relacionadas con los mecanismos de defensa e incremento de los compuestos
antifúngicos y los niveles de compuestos fenólicos (39).
Por otro parte, el Imazalil es un fungicida derivado de los imidazoles. Pertenece
al grupo de los inhibidores de la biosíntesis de los precursores del
ergosterol (IBE), que inhiben la enzima que depende del citocromo P-450, responsable
de la demetilación del ergosterol. Actúa directamente sobre el
patógeno, afecta la permeabilidad celular del hongo y la biosíntesis
de lípidos. Provoca actividad antiesporulante, reducción de la
germinación de esporas y la inflamación de la distorsión
de tubos germinativos y pérdida de citoplasma en células germinadas.
Se ha demostrado la sensibilidad de varios aislados de Colletotrichum
spp. a este fungicida (40, 41).
La aplicación del imazalil + cera y el imazalil + cera + Conserver 21,
(este último es un producto absorbedor del etileno), pudiera modificar
la atmósfera que rodea al fruto, con la consecuente disminución
de la respiración y producción de etileno, así como el
atraso en la maduración (42, 43, 44), que además, da lugar a una
demora en la actividad de diferentes enzimas involucradas en las rutas de biosíntesis
de los compuestos fenólicos (45), como son la polifenoloxidasa o peroxidasa,
principales responsables de la degradación de estos compuestos (46).
Los altos contenidos de fenoles totales en el exocarpio de los frutos de mango
'Super Haden', tratados en poscosecha, están en correspondencia con la
baja incidencia de la antracnosis y la efectividad de estos en su control.
Estos resultados constituyen un aporte al conocimiento sobre el desarrollo y
la severidad de la antracnosis en los frutos de mango 'Super Haden' y la efectividad
de los tratamientos poscosecha para su control, siendo importante determinar
otras enzimas y los procesos relacionados con los frutos y la manifestación
de la enfermedad.
CONCLUSIONES
Nota al pie
A MINAGRI. Reporte Estadístico. La Habana, Cuba, 2010.
B Brecht,
J.; Sargent, S.; Kader, A.; Mitcham, E.; Maul, F.; Brecht, P. y Menocal, O.
Manual práctico para el mejoramiento poscosecha del mango. Inst.
Copyright National Mango Board, Orlando, Florida, 2010, pp. 5-67.
BIBLIOGRAFÍA
1. Pinelo, M.;
Rubilar, M.; Sineiro, J. y Núñez, M. J. ‘‘Extraction of antioxidant
phenolics from almond hulls (Prunus amygdalus) and pine sawdust (Pinus pinaster)’’.
Food Chemistry, vol. 85, no. 2, abril de 2004, pp. 267-273, ISSN 0308-8146,
DOI 10.1016/j.foodchem.2003.06.020.
2. Berardini, N.; Fezer, R.; Conrad, J.; Beifuss, U.; Carle, R. y Schieber,
A. ‘‘Screening of Mango (Mangifera indica L.) Cultivars for Their Contents
of Flavonol O- and Xanthone C-Glycosides, Anthocyanins, and Pectin’’. Journal
of Agricultural and Food Chemistry, vol. 53, no. 5, 1 de marzo de 2005,
pp. 1563-1570, ISSN 0021-8561, DOI 10.1021/jf0484069.
3. Loon, L. C. van; Rep, M. y Pieterse, C. M. J. ‘‘Significance of Inducible
Defense-related Proteins in Infected Plants’’. Annual Review of Phytopathology,
vol. 44, no. 1, 2006, pp. 135-162, ISSN 0066-4286, 0066-4286, DOI 10.1146/annurev.phyto.44.070505.143425.
4. del Río, J. A.; Gómez, P.; Baidez, A. G.; Arcas, M. C.; Botía,
J. M. y Ortuño, A. ‘‘Changes in the Levels of Polymethoxyflavones and
Flavanones as Part of the Defense Mechanism of Citrus sinensis (Cv.
Valencia Late) Fruits against Phytophthora citrophthora’’. Journal
of Agricultural and Food Chemistry, vol. 52, no. 7, 1 de abril de 2004,
pp. 1913-1917, ISSN 0021-8561, DOI 10.1021/jf035038k.
5. Kim, H. G.; Kim, G.-S.; Lee, J. H.; Park, S.; Jeong, W. Y.; Kim, Y.-H.; Kim,
J. H.; Kim, S. T.; Cho, Y. A.; Lee, W. S.; Lee, S. J.; Jin, J. S. y Shin, S.
C. ‘‘Determination of the change of flavonoid components as the defense materials
of Citrus unshiu Marc. fruit peel against Penicillium digitatum
by liquid chromatography coupled with tandem mass spectrometry’’. Food Chemistry,
vol. 128, no. 1, 1 de septiembre de 2011, pp. 49-54, ISSN 0308-8146, DOI 10.1016/j.foodchem.2011.02.075.
6. Miles, T. D.; Vandervoort, C.; Nair, M. G. y Schilder, A. C. ‘‘Characterization
and biological activity of flavonoids from ripe fruit of an anthracnose-resistant
blueberry cultivar’’. Physiological and Molecular Plant Pathology,
vol. 83, julio de 2013, pp. 8-16, ISSN 0885-5765, DOI 10.1016/j.pmpp.2013.02.004.
7. Jeong, S. W.; Kim, H. G.; Park, S.; Lee, J. H.; Kim, Y. H.; Kim, G. S.; Jin,
J. S.; Kwak, Y. S.; Huh, M. R.; Lee, J. E.; Song, Y. y Shin, S. C. ‘‘Variation
in flavonoid levels in Citrus benikoji Hort. ex. Tan. infected by Colletotrichum
gloeosporioides’’. Food Chemistry, vol. 148, 1 de abril de 2014,
pp. 284-288, ISSN 0308-8146, DOI 10.1016/j.foodchem.2013.10.070.
8. Ruiz, G. Y. y Gómez, P. E. ‘‘Elicitors: A Tool for Improving Fruit
Phenolic Content’’. Agriculture, vol. 3, no. 1, 25 de enero de 2013,
pp. 33-52, ISSN 2077-0472, DOI 10.3390/agriculture3010033.
9. Becatti, E.; Chkaiban, L.; Tonutti, P.; Forcato, C.; Bonghi, C. y Ranieri,
A. M. ‘‘Short-Term Postharvest Carbon Dioxide Treatments Induce Selective Molecular
and Metabolic Changes in Grape Berries’’. Journal of Agricultural and Food
Chemistry, vol. 58, no. 13, 14 de julio de 2010, pp. 8012-8020, ISSN 0021-8561,
DOI 10.1021/jf100936x.
10. Crifò, T.; Puglisi, I.; Petrone, G.; Recupero, G. R. y Lo Piero,
A. R. ‘‘Expression analysis in response to low temperature stress in blood oranges:
Implication of the flavonoid biosynthetic pathway’’. Gene, vol. 476,
no. 1–2, 1 de mayo de 2011, pp. 1-9, ISSN 0378-1119, DOI 10.1016/j.gene.2011.02.005.
11. Eichholz, I.; Huyskens, K. S.; Keller, A.; Ulrich, D.; Kroh, L. W. y Rohn,
S. ‘‘UV-B-induced changes of volatile metabolites and phenolic compounds in
blueberries (Vaccinium corymbosum L.)’’. Food Chemistry, vol.
126, no. 1, 1 de mayo de 2011, pp. 60-64, ISSN 0308-8146, DOI 10.1016/j.foodchem.2010.10.071.
12. Danner, M. A.; Sasso, S. A. Z.; Medeiros, J. G. S.; Marchese, J. A. y Mazaro,
S. M. ‘‘Indução de resistência à podridão-parda
em pêssegos pelo uso de eliciadores em pós-colheita’’. Pesquisa
Agropecuária Brasileira, vol. 43, no. 7, julio de 2008, pp. 793-799,
ISSN 0100-204X, DOI 10.1590/S0100-204X2008000700002.
13. Wang, F.; Feng, G. y Chen, K. ‘‘Defense responses of harvested tomato fruit
to burdock fructooligosaccharide, a novel potential elicitor’’. Postharvest
Biology and Technology, vol. 52, no. 1, abril de 2009, pp. 110-116, ISSN
0925-5214, DOI 10.1016/jpostharvbio.2008.09.002.
14. Pan, Y. G. y Liu, X. H. ‘‘Effect of benzo-thiadiazole-7-carbothioic acid
S-methyl ester (BTH) treatment on the resistant substance in postharvest mango
fruits of different varieties’’. African Journal of Biotechnology,
vol. 10, no. 69, 2011, pp. 15521–15528, ISSN 1684-5315, DOI http://dx.doi.org/10.5897/AJB11.2150.
15. Li, M.; Yu, M. L.; Zhang, Z. Q.; Liu, Z. G. y Pan, Y. ‘‘Control of black
spot disease caused by Alternaria alternata on jujube (Ziziphus
jujuba Mill. cv. Dongzao) using HarpinXoo protein’’. The
Journal of Horticultural Science and Biotechnology, vol. 87, no. 3, 1 de
enero de 2012, pp. 250-254, ISSN 1462-0316, DOI 10.1080/14620316.2012.11512860.
16. Tian, S.; Wan, Y.; Qin, G. y Xu, Y. ‘‘Induction of defense responses against
Alternaria rot by different elicitors in harvested pear fruit’’. Applied
Microbiology and Biotechnology, vol. 70, no. 6, 13 de septiembre de 2005,
pp. 729-734, ISSN 0175-7598, 1432-0614, DOI 10.1007/s00253-005-0125-4.
17. Rodríguez, L. E. S.; González, P. J. M. y Mayek, P. N. ‘‘La
Infección de Colletotrichum gloeosporioides (Penz.) Penz. y
Sacc. en Aguacatero (Persea americana Mill.): Aspectos Bioquímicos
y Genéticos’’. Revista Mexicana de Fitopatología, vol.
27, no. 1, enero de 2009, pp. 53-63, ISSN 0185-3309.
18. Mena, N. G.; Valencia, D. T. G.; Piña, G. A. B.; Villanueva, A. R.;
Durán, P. E. y Robles, M. F. ‘‘Degradation capacity of fungi (Colletotrichum
sp., Penicillium sp. and Rhizopus sp.) on mangoes and oranges’’.
African Journal of Agricultural Research, vol. 7, no. 32, 2012, pp. 4564–4574,
ISSN 1991-637X.
19. Kamle, M.; Kumar, P.; Gupta, V. K.; Tiwari, A. K.; Misra, A. K. y Pandey,
B. K. ‘‘Identification and phylogenetic correlation among Colletotrichum
gloeosporioides pathogen of anthracnose for mango’’. Biocatalysis and
Agricultural Biotechnology, vol. 2, no. 3, julio de 2013, pp. 285-287,
ISSN 1878-8181, DOI 10.1016/j.bcab.2013.04.001.
20. Onyeani, C. A.; Osunlaj, S. O.; Owuru, O. O. y Sosanya, O. S. ‘‘Mango Fruit
Anthracnose and the Effects on Mango Yield and Market Values in Southwestern
Nigeria’’. Asian Journal of Agricultural Research, vol. 6, no. 4, 1
de abril de 2012, pp. 171-179, ISSN 18191894, DOI 10.3923/ajar.2012.171.179.
21. López, J. M. y Castaño, Z. J. ‘‘Management of mango anthracnose
[Glomerella cingulata (Stoneman) Spauld. & H. Schrenk] in post-harvest’’.
Agronomía, vol. 18, no. 1, 2010, pp. 47-57, ISSN 0568-3076,
CABDirect2.
22. Hernández, A.; Pérez, J.; Bosch, D. y Castro, N. Clasificación
de los suelos de Cuba 2015. edit. Ediciones INCA, Mayabeque, Cuba, 2015,
93 p.,
ISBN 978-959-7023-77-7.
23. Ploetz, R. C.; Zentmyer, G. A.; Nishijima, W. T.; Rohrbach, K. G. y Ohr,
H. D. Compendium of tropical fruit diseases. [en línea]. vol.
8, edit. American Phytopathological Society (APS), 1994, 88 p., ISBN 0-89054-162-0,
[Consultado: 2 de diciembre de 2015], Disponible en: <http://www.cabdirect.org/abstracts/19942309422.html>.
24. Mulkay, T.; Rodríguez, J.; Cáceres, I. y Paumier, A. ‘‘Caracterización
de los daños por antracnosis en frutos de mango (Mangifera indica
L.) a nivel de macro y ultraestructural’’. Citrifrut, vol. 22, no.
1-2-3, 2005, pp. 25-28, ISSN 1607-5072.
25. Singleton, V. L. y Rossi, J. A. ‘‘Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic
Acid Reagents’’. American Journal of Enology and Viticulture, vol.
16, no. 3, 1 de enero de 1965, pp. 144-158, ISSN 0002-9254.
26. Abbott, W. S. ‘‘A Method of Computing the Effectiveness of an Insecticide’’.
Journal of Economic Entomology, vol. 18, no. 2, 1 de abril de 1925,
pp. 265-267, ISSN 0022-0493, 1938-291X, DOI 10.1093/jee/18.2.265a.
27. Akem, C. N. ‘‘Mango Anthracnose Disease: Present Status and Future Research
Priorities’’. Plant Pathology Journal, vol. 5, no. 3, 1 de marzo de
2006, pp. 266-273, ISSN 18125387, 18125425, DOI 10.3923/ppj.2006.266.273.
28. Pandey, A.; Yadava, L. P.; Mishra, R. K.; Pandey, B. K.; Muthukumar, M.
y Chauhan, U. K. ‘‘Studies on the incident and pathogenesis of Colletotrichum
gloeosporioides penz. causes anthracnose of mango’’. International
Journal of Science and Nature, vol. 3, no. 2, 2012, pp. 220–232, ISSN 2229-6441,
0973-3140.
29. Cantos, E.; Espín, J. C.; Fernández, M. J.; Oliva, J. y Tomás-Barberán,
F. A. ‘‘Postharvest UV-C-Irradiated Grapes as a Potential Source for Producing
Stilbene-Enriched Red Wines’’. Journal of Agricultural and Food Chemistry,
vol. 51, no. 5, 1 de febrero de 2003, pp. 1208-1214, ISSN 0021-8561, DOI 10.1021/jf020939z.
30. Kumah, P.; Appiah, F. y Opoku, D. J. K. ‘‘Effect of hot water treatment
on quality and shelf-life of Keitt mango’’. Agriculture and Biology Journal
of North America, vol. 2, no. 5, 2011, pp. 806–817, ISSN 2151-7517, 2151-
7525, DOI 10.5251/abjna.2011.2.5.806.817.
31. Santamaría, B. F.; Díaz, P. R.; Gutiérrez, A. O.; Santamaría,
F. J. y Larqué, S. A. ‘‘Control de dos especies de Colletotrichum
causantes de antracnosis en frutos de papaya Maradol’’. Revista Mexicana
de Ciencias Agrícolas, vol. 2, no. 5, octubre de 2011, pp. 631-643,
ISSN 2007-0934.
32. Lado, J.; Pérez, F. E.; De Nigris, A.; Dol, I. y Knochen, M. ‘‘Residuos
en frutos de naranja del fungicida imazalil aplicado en postcosecha y su efecto
en el control de moho verde’’. Agrociencia Uruguay, vol. 17, no. 2,
diciembre de 2013, pp. 83-90, ISSN 2301-1548.
33. Talcott, S. T.; Moore, J. P.; Lounds, S. A. J. y Percival, S. S. ‘‘Ripening
Associated Phytochemical Changes in Mangos (Mangifera indica) Following
Thermal Quarantine and Low-Temperature Storage’’. Journal of Food Science,
vol. 70, no. 5, 1 de junio de 2005, pp. C337-C341, ISSN 1750-3841, DOI 10.1111/j.1365-2621.2005.tb09963.x.
34. Gonzalez, A. G. A.; Villa, R. J. A.; Ayala, Z. J. F. y Yahia, E. M. ‘‘Improvement
of the antioxidant status of tropical fruits as a secondary response to some
postharvest treatments’’. Trends in Food Science & Technology,
vol. 21, no. 10, octubre de 2010, pp. 475-482, ISSN 0924-2244, DOI 10.1016/j.tifs.2010.07.004.
35. Ummarat, N.; Matsumoto, T. K.; Wall, M. M. y Seraypheap, K. ‘‘Changes in
antioxidants and fruit quality in hot water-treated ‘Hom Thong’ banana fruit
during storage’’. Scientia Horticulturae, vol. 130, no. 4, 31 de octubre
de 2011, pp. 801-807, ISSN 0304-4238, DOI 10.1016/j.scienta.2011.09.006.
36. Liu, J.; Sui, Y.; Wisniewski, M.; Droby, S.; Tian, S.; Norelli, J. y Hershkovitz,
V. ‘‘Effect of heat treatment on inhibition of Monilinia fructicola
and induction of disease resistance in peach fruit’’. Postharvest Biology
and Technology, vol. 65, marzo de 2012, pp. 61-68, ISSN 0925-5214, DOI
10.1016/j.postharvbio.2011.11.002.
37. Escribano, S. y Mitcham, E. J. ‘‘Progress in heat treatments’’. Stewart
Postharvest Review, vol. 10, no. 3, 1 de diciembre de 2014, pp. 1-6, ISSN
1745-9656.
38. Li, X.; Zhu, X.; Zhao, N.; Fu, D.; Li, J.; Chen, W. y Chen, W. ‘‘Effects
of hot water treatment on anthracnose disease in papaya fruit and its possible
mechanism’’. Postharvest Biology and Technology, vol. 86, diciembre
de 2013, pp. 437-446, ISSN 0925-5214, DOI 10.1016/j.postharvbio.2013.07.037.
39. Yuan, L.; Bi, Y.; Ge, Y.; Wang, Y.; Liu, Y. y Li, G. ‘‘Postharvest hot water
dipping reduces decay by inducing disease resistance and maintaining firmness
in muskmelon (Cucumis melo L.) fruit’’. Scientia Horticulturae,
vol. 161, 24 de septiembre de 2013, pp. 101-110, ISSN 0304-4238, DOI 10.1016/j.scienta.2013.06.041.
40. Sombardier, A.; Dufour, M.-C.; Blancard, D. y Corio, C. M.-F. ‘‘Sensitivity
of Podosphaera aphanis isolates to DMI fungicides: distribution and
reduced cross-sensitivity’’. Pest Management Science, vol. 66, no.
1, 1 de enero de 2010, pp. 35-43, ISSN 1526-4998, DOI 10.1002/ps.1827.
41. Salazar, E.; Hernández, R.; Tapia, A. y Gómez, A. L. ‘‘Identificación
molecular del hongo «Colletotrichum» spp. aislado de banano
(«Musa» spp.) de altura en la zona de Turrialba y determinación
de su sensibilidad a fungicidas poscosecha’’. Agronomía costarricense:
Revista de Ciencias Agrícolas, vol. 36, no. 1, 2012, pp. 53-68,
ISSN 0377-9424.
42. Guillén, F.; Garrido, D. V.; Coll, P. J. Z.; Serrano, M. S.; Valverde,
J. M.; Mula, H. M. D.; García, S. C. y Romero, D. M. ‘‘Desarrollo de
un envase activo para mejorar la calidad de ciruela y mantener sus propiedades
antioxidantes’’. Horticultura Global, no. 305, 2012, pp. 60-66, ISSN
2173-5042.
43. Zhang, Z.; Tian, S.; Zhu, Z.; Xu, Y. y Qin, G. ‘‘Effects of 1-methylcyclopropene
(1-MCP) on ripening and resistance of jujube (Zizyphus jujuba cv. Huping)
fruit against postharvest disease’’. LWT - Food Science and Technology,
vol. 45, no. 1, enero de 2012, pp. 13-19, ISSN 0023-6438, DOI 10.1016/j.lwt.2011.07.030.
44. González, E. M.; Fernández, A. E. L.; Páez, H. R.;
Oca, M. M. M. de y Gómez, B. T. ‘‘Uso combinado de 1-Meticiclopropeno
y emulsiones de cera en la conservación de guanábana (Annona
muricata)’’. Revista Brasileira de Fruticultura, vol. 36, no.
SPE1, 2014, pp. 296-304, ISSN 0100-2945, DOI 10.1590/S0100-29452014000500035.
45. Desjardins, Y. ‘‘Physiological and ecological functions and biosynthesis
of health-promoting compounds in fruit and vegetables’’. En: Tomás B.
F. A. y Gil M. I., Improving the Health-Promoting Properties of Fruit and
Vegetable Products, edit. Elsevier, USA, 23 de abril de 2008, pp. 201–247,
ISBN 978-1-84569-428-9.
46. Pourcel, L.; Routaboul, J.-M.; Cheynier, V.; Lepiniec, L. y Debeaujon, I.
‘‘Flavonoid oxidation in plants: from biochemical properties to physiological
functions’’. Trends in Plant Science, vol. 12, no. 1, 1 de enero de
2007, pp. 29-36, ISSN 1360-1385, DOI 10.1016/j.tplants.2006.11.006.
Recibido: 13 de
diciembre de 2014
Aceptado: 2 de septiembre de 2015
Akira Pérez Márquez, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700. Email: akiraperez82@gmail.com