Efecto del déficit hídrico sobre cambios morfo-fisiológicos y bioquímicos en plantas micropropagadas de piña ‘MD-2’ en la etapa final de aclimatización
Water effect deficit on morpho-physiological and biochemical changes in ‘MD-2’ micro-propagated pineapple plants at the end of acclimatization stage
René C. Rodríguez-Escriba,I Ibraín D. Rodríguez-Cartaya,II Gustavo Y. Lorente,I Dariel López,I Roberto E. Izquierdo,I Lucía S. Borroto,I Camilo Bonet,III Yolanda Garza-García,IV Carlos E. Aragón,I Florencio E. Podestá,V Romelio Rodríguez,I Justo L. González-OlmedoI
ICentro
de Bioplantas. Universidad de Ciego de Ávila. Ciego de Ávila,
Cuba.
IIUniversidad de La Habana. La Habana, Cuba.
IIIInstituto de investigaciones de Ingeniería Agrícola,
La Habana, Cuba
IVUniversidad Autónoma de Coahuila, México.
VCentro de Estudios Fotosintéticos y Bioquímicos. Fac.
de Ciencias Bioquímicas y Farmacéuticas. Universidad Nacional
de Rosario. Rosario, Argentina.
RESUMEN
La tecnología actual de micropropagación de piña presenta problemas con la transición de las plantas al campo, dichos problemas se asocian al drástico cambio de las condiciones ambientales unido al pobre endurecimiento de las plantas para su tránsito. Una posibilidad de su preparación es la inducción de mecanismos defensivos mediante el déficit hídrico y consigo la modulación de CAM (Metabolismo Ácido de las Crasuláceas, siglas en inglés) por lo que en este trabajo se estudiaron plantas crecidas durante 30 días en dos regímenes hídricos (plantas con riego y sin riego), después de este momento ambos grupos se regaron hasta la saturación del sustrato durante 15 días. Las determinaciones se realizaron en la hoja “D” con frecuencia quincenal. Se determinó el intercambio gaseoso, eficiencia en el uso del agua (EUA), porcentaje de captación de CO2 en la noche, contenido de clorofilas, niveles de ácidos orgánicos, índice de suculencia (IS) y actividad de superóxido dismutasa (SOD) (EC 1.15.1.1). A los 15 días las plantas sin riego tuvieron la mejor respuesta para su traslado a campo, con el aumento de la expresión de CAM, sustentado por la disminución del contenido de clorofilas totales y por el incremento del porcentaje de asimilación de CO2 durante la noche, EUA, IS y actividad SOD. A los 30 días las plantas sin riego tuvieron una respuesta CAM de mayor intensidad, con el 100 % de captación de CO2 durante la noche, pero su rápida recuperación con el establecimiento del riego hasta la saturación del sustrato, lo cual demostró alta tolerancia a la sequía y elevada plasticidad metabólica.
Palabras clave: ciclo CAM, fotosíntesis, vitroplantas.
ABSTRACT
Current technology of pineapple micro-propagation has problems with the plants transition to field, these problems are associated to drastic changes of environmental conditions linked to poor hardening plant for such transit. A possibility of preparation is the induction of defense mechanisms to drought stress and get modulating CAM, so that the object of this paper was study plants grown for 30 days in two water status (well-watered plants and non-watered plants) after 30 days both groups were full-watered during 15 days. Measurements were made on D-leaf, in which were determinates the gas exchange, water-efficiency use (WEU), night CO2 uptake percentage, chlorophyll content, organic acids levels, succulence index (IS) and superoxide dismutase activity (SOD) (EC 1.15.1.1). After 15 days of drought, plants had the best response to field transfer, with increase in CAM expression, supported by the decline in total chlorophyll content and increases in the night CO2 uptake percentage, WEU, SI, and SOD activity. After 30 days of drought plants had a CAM strong response, with 100 % of CO2 uptake during night, but its rapid recovery with the establishment of irrigation to saturation of the substrate, showing high drought tolerance and great metabolic plasticity.
Key words: CAM cycle, photosynthesis, vitroplants.
INTRODUCCIÓN
El híbrido
‘MD-2’ posee caracteres de elevada importancia agronómica con altos rendimientos
productivos y calidad de fruta comparada con el resto de los cultivares de piña
(1).
La necesidad de introducir ‘MD-2’ en las plantaciones cubanas, requiere de grandes
producciones de material de propagación en el menor tiempo posible, lo
cual es posible con la utilización de las técnicas de cultivo
in vitro como herramientas de la micropropagación, tecnología
que permite obtener gran número de plantas en poco tiempo y con mejor
calidad; sin embargo, los laboratorios de micropropagación tienen limitaciones,
dicho proceso tecnológico induce en las plantas un estado fisiológico
desventajoso para la transición a condiciones ex vitro y, como
consecuencia, la obtención de bajas tasas de crecimiento e insuficientes
porcentajes de supervivencia durante la aclimatización (2).
Se han realizado investigaciones para determinar el mejor medio químico
o físico de cultivo in vitro para obtener plantas capaces de
aclimatarse a las condiciones ex vitro (3, 4) y en la fase de aclimatización
(5, 6). Sin embargo, el protocolo utilizado para la aclimatización presenta
problemas con la transición de las plantas al campo, una vez que son
plantadas muestran quemaduras en las hojas y senescencia en algunos tejidos
con una consecuente baja supervivencia.
Estos problemas están asociados al estrés provocado por el drástico
cambio entre las condiciones ambientales de la fase final de aclimatización
y el campo (7), por lo que es necesario desarrollar investigaciones encaminadas
a disminuir el efecto negativo que ejerce el ambiente en las plantas de piña
‘MD-2’ en la transición hacia el campo.
Ananas comosus presenta metabolismo ácido de las crasuláceas
(CAM). Las plantas con CAM pueden operar de diferentes modos: “CAM constitutiva”
o “CAM obligada”, con alta acumulación de ácidos (ΔH+)
y captación de CO2 durante la noche; “CAM inducible o “CAM
facultativa”, conocidas también como “C3-CAM” con una forma C3 de fijación
de CO2 y variación nula de ΔH+ en el estado
no inducido; pero en el estado inducido presenta pequeñas fijaciones
de CO2 en el día y altos niveles de ΔH+ durante
la noche; “CAM cíclica” con fijación de CO2 durante
el día y ΔH+ pero no presenta apertura estomática
durante la noche; CAM “Idling” o “CAM fútil”, con cierre estomático
y pequeñas variaciones de ΔH+ durante todo el día
y la noche en plantas severamente estresadas (8).
Estos comportamientos pueden exhibir diferente fisiotipos dentro del propio
metabolismo, como en el caso de las plantas “CAM constitutivas”, las cuales
pueden ser fuertes o débiles, de acuerdo a la magnitud de expresión
de la ruta C4. La piña en su hábitat o en condiciones de producción
en el campo responde como una “CAM constitutiva fuerte” (9), pero se ha demostrado
que las plantas micropropagadas pueden exhibir un fisiotipo diferente, estas
pueden ser C3 o CAM de acuerdo a las condiciones de crecimiento in vitro
(3, 4, 10, 11); además, estudios previos demostraron que un incremento
de la intensidad lumínica aumenta la magnitud de expresión CAM
en la etapa de aclimatización y que las plantas tratadas adquirieron
un estado morfo-fisiológico y bioquímico ventajoso para la transición
al campo (1).
Teniendo en cuenta que CAM se expresa y modula con cambios en las variables
ambientales como intensidad lumínica, humedad, nutrición y temperatura
(12), desde el punto de vista práctico y con una perspectiva económica
se pueden realizar manejos con frecuencia e intensidad de riego para inducir
una mejor respuesta morfo-fisiológica y bioquímica de las plantas
en la etapa final de aclimatización favoreciendo el tránsito aclimatización-campo
y por ende el incremento de la supervivencia.
MATERIALES Y MÉTODOS
Esta investigación
se realizó entre los meses septiembre y noviembre del 2014 en el área
de aclimatización del Centro de Bioplantas. Se utilizaron plantas de
piña Ananas comosus (L.) Merr. ‘MD-2’ micropropagadas, según
el protocolo establecido en el centro de Bioplantas. Se seleccionaron plantas
de más de 4,0 cm de longitud, con 7,0 a 8,0 g de masa fresca y cinco
y seis hojas funcionales y se sumergieron durante cinco minutos en una solución
de Previcur Energy® (Bayer CropScience) [3,0 mL L-1]
(13). Seguidamente se plantaron en envases plásticos con un volumen de
sustrato de 256,26 cm3 con relación 1:1 (v/v) entre suelo
ferralítico rojo y cachaza (derivado de la caña de azúcar)
(6).
Las plantas se aclimatizaron en una casa de cultivo con condiciones promedios
de 80±3 % de humedad relativa, 25,5±2 °C de temperatura (determinados
con un termo-higrómetro digital “TECPEL® modelo DTM- 303”),
luz y fotoperiodo natural con un Flujo de Fotones Fotosintéticos (FFF)
de 400 ± 25 µmol m-2s-1 y condiciones atmosféricas
de concentración de CO2 entre 330 y 375 µmol mol-1
(determinados con un sistema portátil de fotosíntesis “PP Systems
CIRAS-2 Portable Photosynthesis System”). El riego se realizó por aspersión
durante 10 minutos a las 9:00 y 14:00 h diariamente. Se aplicó fertilización
foliar con una mochila de fumigación cada 15 días con NPK cristalino
[1,0 g L-1] y Multimicro Combi (Haifa Chemicals Ltd., Haifa Bay 26120,
Israel) [62,5 mg L-1].
Después de cinco meses en aclimatización se seleccionaron 400
plantas de 34-36 g de masa fresca con 11-12 hojas y 12-13 raíces funcionales
y se trasladaron al exterior de la casa de cultivo con una intensidad lumínica
de 800 µmol m-2s-1. Los grupos se sometieron a diferentes
condiciones de riego según Tabla I.
Se aplicó
fertilización foliar a las 9:00 h en el momento inicial del experimento
(N-P-K cristalino [1,0 g L-1] y Multimicro Combi (Haifa Chemicals
Ltd., Haifa Bay 26120, Israel) [62,5 mg L-1]). El día 31 del
experimento se aplicó fertilización foliar a las 9:00 h (N-P-K
cristalino [1,0 g L-1] y Multimicro Combi (Haifa Chemicals Ltd.,
Haifa Bay 26120, Israel) [62,5 mg L-1]). El riego se realizó
con una regadera manual, distribuyendo de manera homogénea 100 L de agua
sobre la superficie foliar de las plantas cada 24 h.
Determinaciones morfológicas: se tomaron 30 plantas para la
determinación del número de hojas y longitud y ancho de la hoja
D. Además se determinó la masa fresca y seca de 10 discos de hojas
de 0,785 cm2 en cuatro hojas D colectadas a las 15:00 h y se calculó
el contenido de agua.
Cuantificación del intercambio gaseoso en las hojas: se cuantificó
el intercambio gaseoso cada tres horas hasta completar un día. Se utilizó
un analizador infrarrojo de gases (IRGA) como parte de un sistema de fotosíntesis
portátil (PP Systems CIRAS-2 Portable Photosynthesis System). Se tomaron
tres hojas D y se realizaron 30 mediciones para cada momento y tratamiento.
Las determinaciones se realizaron en condiciones de temperatura y humedad relativa
ambientales, con un FFF de 600 µmol m-2s-1 (14)
. Para el análisis de los resultados se tomó el promedio entre
las 21:00 y las 9:00 h. Se calculó la eficiencia en el uso del agua con
la división de la tasa de asimilación de CO2 y la tasa
transpiratoria. Además se determinó el porcentaje que representó
la asimilación de CO2 entre dichas horas respecto al día
completo.
Cuantificación del contenido de clorofilas: se colectaron tres hojas
D a las 15:00 h y se conservaron en nitrógeno líquido (NL) hasta
el momento de las determinaciones. Se tomó 0,1 g de hojas y se maceraron
en un mortero con NL hasta obtener un polvo fino. Se adicionaron 500 µL
de acetona al 80 % con 2,5 mM de Fosfato de sodio pH 7,8, posteriormente se
centrifugó a 12 000 g durante 15 minutos a 4 ºC con una centrífuga
refrigerada al vacío Beckman (JB-21). Se colectó el sobrenadante
y se midió la absorbancia a 664 nm y 647 nm. Se calcularon las concentraciones
de clorofilas a, b y totales (15). Se utilizó un espectrofotómetro
“Pharmacia Bio-Spectrophotometer”. Los resultados se expresaron en función
de la masa fresca y se convirtieron de µg Cfls g-1 MF a µg
Cfls cm-2 como es descrito (3).
Cálculo del índice de suculencia: el Índice de
Suculencia (IS) se calculó utilizando los valores del contenido de clorofilas
totales [Cfls (a+b)] y el contenido de agua (CA) de los discos de hojas “D”.
Se utilizó la ecuación IS= CA [Cfls (a+b)]-1 (16).
Determinación de los niveles de ácidos orgánicos:
se tomó 1g de hojas, se adicionaron 10 mL de Etanol 50 % y se incubaron
en un baño a 90 ºC durante 20 minutos. Se separó la fase
líquida y se realizó una valoración ácido-base,
con NaOH [20 mM] y fenolftaleína [1 mg L-1] como indicador
(17). Los resultados se expresaron en µmol H+ h-1g-1
MF.
Cuantificación de proteínas y actividad de la enzima Superóxido
Dismutasa: se pesaron 0,5 g de material vegetal y se maceraron con NL hasta
obtener un polvo fino, se añadieron 2 mL de buffer fosfato de potasio
50 mM pH 7,8 con 1 mM EDTA; 0,1 % Triton X-100 (v/v); 1 % PVP; 1 mM DTT y 1
mM PMSF), luego se centrifugó a 15 000 g durante 20 minutos a 4 ºC
con una centrífuga refrigerada al vacío Beckman (JB-21) (16).
Se colectó el sobrenadante y se cuantificó el contenido de proteínas
(18), utilizando una curva patrón con “Bobine Sero Albumin” (BSA) como
proteína estándar. El contenido de proteínas se expresó
en mg Prot. g-1 MS. La actividad de la enzima Superóxido dismutasa
(SOD) (EC 1.15.1.1) se determinó por el método de oxidorreducción
del citocromo C como es descrito (16). La actividad enzimática se expresó
como U mg-1 MS. Una unidad de actividad SOD se definió como
la cantidad de enzima capaz de inhibir el 50 % de velocidad de reducción
del citocromo C.
Análisis estadístico: todas las determinaciones fisiológicas
y bioquímicas se replicaron tres veces en tres muestras biológicas
diferentes para cada momento de evaluación y tratamiento. Se utilizó
el software Statgraphics Centurion XV (19). Los resultados se procesaron con
“ANOVA factorial” seguido por “Tukey’s Multiple Range test” para rangos múltiples,
o “Kruskal-Wallis” seguido por “Student-Newman-Keuls” en los casos necesarios.
Todas las pruebas estadísticas fueron realizadas con un nivel de significación
de un 5 %.
RESULTADOS Y DISCUSIÓN
Efecto
del déficit hídrico en indicadores morfológicos
Después de 15 días en las plantas sometidas al déficit
hídrico (SR) se observó un cese de la elongación de la
hoja D (Figura 1A; sin embargo, a partir de los 30 días,
cuando ambos grupos fueron regados hasta la saturación del sustrato se
observó un aumento de la longitud de dicha hoja en este mismo tratamiento.
Por otra parte, durante el periodo de déficit hídrico (hasta 30
días) no se observó emisión de hojas en las plantas SR
(Figura 1C), pero sí
con el restablecimiento del riego.
El déficit
hídrico afectó tanto la elongación de las plantas SR como
la emisión de hojas, lo cual está relacionado con una mejor aclimatación
a condiciones ambientales adversas. La tasa de crecimiento y por ende la productividad
de biomasa de plantas CAM terrestres son muy bajas comparadas con plantas C3
y C4 bajo condiciones poco estresantes; sin embargo, dentro de una misma especie
CAM la productividad de biomasa está favorecida por condiciones ambientales
favorables (12, 20, 21) y viceversa.
En contraste se observó un incremento continuo del ancho de la hoja D
durante el periodo de déficit hídrico (Figura
1B), seguido por un cese de su incremento después del establecimiento
del riego (a los 45 días). Las plantas de piña micropropagadas
en condiciones de aclimatización presentan una relación directa
entre el aumento del ancho de dicha hoja y la expresión (3) y modulación
(16) de CAM bajo condiciones estresantes.
Es importante señalar que también se observó un incremento
continuo aunque en menor grado en las plantas CR, esto pudo estar asociado al
traslado de las plantas al exterior de la casa de cultivo, unido al incremento
de la edad fisiológica de las plantas. Ambos grupos se expusieron a la
luz solar directa durante 30 días antes de iniciar el experimento, de
esta forma el incremento del ancho de la hoja “D” en las plantas CR pudo estar
relacionado con un aumento gradual de la expresión CAM, como una respuesta
a la alta intensidad lumínica y al incremento de la edad fisiológica
de las plantas. Además, la tasa evapotranspiratoria en el exterior de
la casa de cultivo debió ser superior que en el interior de esta, considerando
que en el exterior existió una menor humedad relativa, mayor velocidad
del viento y altos niveles de irradiación, lo cual favorece la difusión
del agua al aire tanto del suelo como de la superficie foliar, de modo que el
riego establecido en el interior de la casa de cultivo pudo ser insuficiente
en el exterior, y por ende las plantas CR pudieron quedar expuestas a un déficit
hídrico ligero que provocaría el incremento gradual de la magnitud
de expresión CAM, y como respuesta el incremento del ancho de la hoja
“D” durante el periodo experimental.
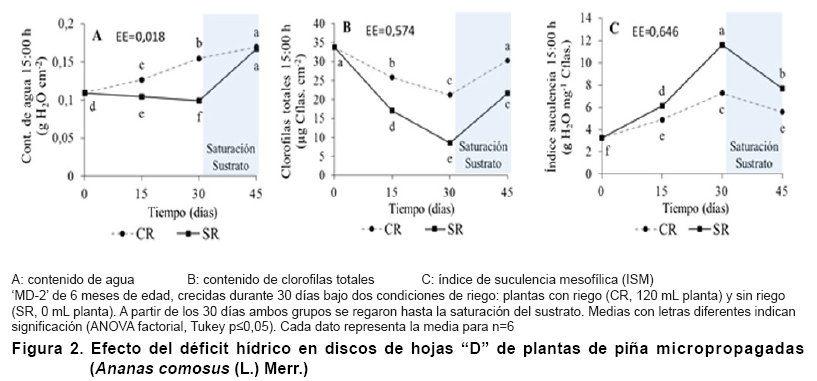
Efecto del déficit hídrico en el contenido de agua, clorofilas
totales e índice de suculencia
El contenido de agua de las hojas en las plantas SR disminuyó durante
el periodo de déficit hídrico en cada momento de evaluación
hasta los 30 días (Figura 2A); sin embargo, cuando
ambos grupos se regaron hasta la saturación del sustrato se observó
un incremento considerable del contenido de agua de las hojas en las plantas
SR (45 días), en este momento el contenido de agua de las hojas de las
plantas SR alcanzó valores similares a las plantas CR.
Esto pudo estar
asociado a la síntesis de tejidos acuíferos en las plantas SR
durante el periodo de déficit hídrico, o a un incremento del potencial
osmótico de las células asociado al influjo de carbohidratos y
ácido málico hacia el interior del tonoplasto.
Por otra parte en el contenido de clorofilas totales se observó una tendencia
similar en ambos grupos (Figura
2B), con una disminución continua hasta los 30 días, seguido
de un incremento con el establecimiento del riego diario hasta la saturación
del sustrato (45 días). Sin embargo, las plantas CR presentaron valores
superiores a las plantas SR en todos los momentos de evaluación.
En el caso del índice de suculencia mesofílica ambos grupos presentaron
una tendencia similar durante todo el experimento pero un mayor grado en las
plantas SR (Figura 2C),
con un aumento de la suculencia de las hojas durante los primeros 30 días
y una disminución con el establecimiento del riego hasta la saturación
del sustrato.
El índice de suculencia mesofílica representa la relación
entre los tejidos acuíferos y los tejidos fotosintéticos.
En este experimento se observó una disminución del contenido de
agua en las hojas de las plantas SR provocado por el déficit hídrico
severo; sin embargo, también se observó una disminución
de los contenidos de clorofilas totales, esto indica que el déficit hídrico
provocó una disminución del contenido de clorofilas totales en
mayor grado que la disminución del contenido de agua de las hojas, lo
cual e confiere mayor suculencia y magnitud de expresión CAM a las plantas
SR.
Una planta es considerada CAM cuando este índice es superior a la unidad
(8), y a mayor suculencia de tallos y hojas mayor magnitud de expresión
CAM y mejor aclimatación a condiciones ambientales adversas (22, 23);
sin embargo, el IS también se incrementó en las plantas CR durante
el periodo de déficit hídrico, lo cual sustenta la conjetura de
que en este grupo existió un aumento gradual de la magnitud de expresión
CAM, como una respuesta al aumento de la edad fisiológica y el traslado
de las plantas al exterior de la casa de cultivo.
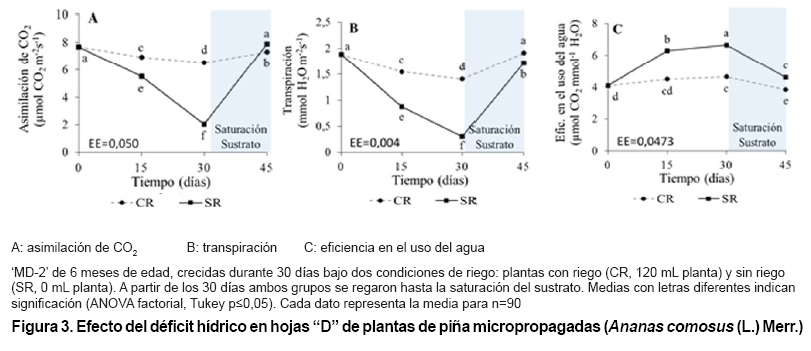
Efecto del déficit hídrico en el intercambio gaseoso
Las plantas SR tuvieron una disminución de la asimilación de CO2
a los 15 y 30 días de déficit hídrico respecto a las plantas
CR y al momento inicial del experimento (Figura 3A). El establecimiento
del riego hasta la saturación del sustrato favoreció la captación
durante la noche en este mismo grupo.
Se observó
una disminución continua de la transpiración en las plantas SR
durante todos los momentos de evaluaciones en el periodo de déficit hídrico,
con diferencias significativas entre ellos y entre tratamientos. En el establecimiento
del riego hasta la saturación del sustrato se observó un incremento
considerable de la transpiración en este mismo grupo de plantas, alcanzando
valores cercanos a las plantas CR aunque estadísticamente diferentes.
Las plantas SR tuvieron un incremento de la eficiencia del uso del agua durante
el periodo de déficit hídrico, con diferencias significativas
respecto a las plantas CR en todos los momentos evaluados; sin embargo ,con
el restablecimiento del riego se observó una disminución significativa
de este indicador en ambos grupos pero con valores superiores en las plantas
SR.
El aumento de la magnitud de expresión CAM está relacionado con
el aumento del porcentaje de captación de CO2 durante la noche
(24), para las plantas CAM constituye un beneficio captar mayor cantidad de
CO2 en momentos donde las condiciones ambientales son menos drásticas,
y de esta forma evitar una pérdida excesiva de agua (12, 25, 26).
El déficit hídrico provocó una disminución de la
captación de CO2 de las plantas SR; sin embargo, el porcentaje
de asimilación de CO2 entre las 21:00 y 9:00 h se incrementó
continuamente en las plantas SR (0-30 días) durante el periodo de déficit
hídrico (Tabla II).
Cuando las plantas
CAM se exponen a déficit hídrico presentan una disminución
de la captación de CO2 durante el día, sobre todo en
horas cercanas al amanecer y anochecer, pero cuando el déficit hídrico
es prolongado o severo cesan la captación de CO2 durante todo
o la mayor parte del ciclo diario (17, 27, 28, 29), en este estado las plantas
CAM para su supervivencia captan y por ende reciclan todo o gran parte del CO2
liberado durante la respiración (30, 31), y esta respuesta ante condiciones
desfavorables se denomina “CAM Idling” o “CAM fútil” (8).
Se ha demostrado que en condiciones favorables de crecimiento, el CO2
captado por la piña durante la noche (fase I) representa más del
75 % del CO2 asimilado durante el día completo; sin embargo,
ante el déficit hídrico prolongado y altas temperaturas durante
el día pueden incrementar dicho porcentaje, lo cual le confiere elevada
tolerancia a la sequía (32). Esto explica el incremento del porcentaje
de asimilación de CO2 durante la noche en las plantas SR (Tabla
II) ante el déficit hídrico por 30 días, donde dicho
porcentaje representó aproximadamente el 100 % de la captación
durante todo el día, lo cual demuestra que las plantas SR tuvieron una
respuesta similar a “CAM Idling” ante el déficit hídrico severo,
con cierre estomático durante casi todo el día y apertura de los
mismos solo cuando el gradiente para la transpiración fue mínimo.
Las plantas de piña micropropagadas ante déficit hídrico
severo también respondieron de manera similar a plantas “CAM Idling”,
en lugar de “CAM facultativas” (3, 4, 10, 11), o “CAM constitutivas” (1) como
se ha demostrado para plantas de piña provenientes del cultivo in
vitro. A pesar del déficit hídrico durante 30 días,
15 días después del restablecimiento del riego hasta la saturación
del sustrato las plantas SR presentaron una rápida recuperación,
demostrando una elevada plasticidad metabólica. Además, en este
mismo momento también se observó un aumento en la longitud de
la hoja “D” (Figura 1A)
y número de hojas (Figura
1C) lo cual permite conjeturar que con la rehidratación de los tejidos
de las plantas SR (Figura
2A) la partición de carbohidratos debió favorecer el crecimiento
en mayor proporción.
En el caso de la transpiración las plantas SR presentaron una disminución
de la transpiración con el aumento del déficit hídrico
hasta 30 días (Figura
3B), a partir de este momento se observó un aumento considerable
en la traspiración con el establecimiento del riego.
La disminución significativa de la transpiración en las plantas
SR a los 15 y 30 días sustenta la hipótesis del incremento de
la magnitud de CAM; sin embargo, la disminución considerable a los 30
días está relacionada además con la disminución
de la asimilación de CO2 (Figura
3A) y del contenido de agua de las hojas (Figura
2A), ambos indicadores en estrecha relación con el déficit
hídrico implicando cierre estomático, lo que permite conjeturar
de que a los 30 días las plantas SR respondieron de forma similar a “CAM
Idling”. Además el déficit hídrico incrementó la
eficiencia en el uso del agua durante la noche en las plantas SR a los 15 días
(Figura 3C).
Un incremento en la eficiencia hídrica en las plantas CAM está
relacionado con un incremento de la magnitud de expresión en plantas
CAM terrestres (12). Respecto al tamaño de los órganos de las
plantas CAM, los estomas aparecen en bajas densidades y presentan bajas conductividades
al vapor de agua, reflejando una alta capacidad de almacenaje de agua en las
hojas y tallos, disminución de la relación área-volumen
de las hojas y por ende elevada eficiencia hídrica (12). Esto explica
el incremento del contenido de agua de las hojas en las plantas SR a los 45
días (Figura 2A),
y el aumento de la eficiencia hídrica de las plantas SR a los 15 y 45
días de evaluación, además sustenta la conjetura de que
el incremento considerable del contenido de agua en las plantas SR a los 45
días pudo estar relacionado con la síntesis de tejidos de reserva
de agua (Figura 2A)
unido al incremento del potencial osmótico de las células.
Efecto del déficit hídrico en los niveles de ácidos
orgánicos
Se observó en la Figura 4 una tendencia contraria en
los niveles de ácidos orgánicos de ambos grupos. El déficit
hídrico provocó una disminución de los niveles de ácidos
orgánicos (AO) en las plantas SR hasta los 30 días de sequía,
en contraste los niveles de AO en las plantas CR aumentó durante el periodo
de déficit hídrico y luego disminuyó a los 45 con el establecimiento
del riego hasta la saturación del sustrato.
En las plantas
CAM el CO2 asimilado durante la noche es fijado por la enzima fosfoenolpiruvato
carboxilasa (PEPC) en fosfoenolpiruvato (PEP) el cual es convertido a malato
y transportado al interior de la vacuola donde se almacena en forma de ácido
málico (33); además, los cambios de los niveles de AO se corresponden
fundamentalmente con los niveles ácidos málico, cítrico
e isocítrico; sin embargo, en Ananas comosus los cambios en
los niveles de AO se corresponden fundamentalmente con los niveles de ácido
málico, permaneciendo constantes los niveles de ácido cítrico
y despreciables los de isocítrico (16), además la relación
entre los niveles de AO y málico responde a una estequiometria 2:1 (24).
De esta forma para la piña se puede establecer una relación directa
entre los niveles de AO y los niveles de ácido málico producto
de la asimilación nocturna, y por ende se puede decir que la cantidad
de malato acumulado durante el periodo de déficit hídrico en las
plantas SR fue inferior a los niveles de las plantas CR, lo cual sustenta la
disminución de la asimilación nocturna de CO2 (Figura
3A), la disminución de la rapidez de elongación de la hoja
D y la cesación de la emisión de hojas durante el periodo de déficit
hídrico.
El incremento de los niveles de ácidos orgánicos a los 45 días
alcanzó valores similares en ambos grupos a los 15 días de estudio,
esto implica una recuperación de las plantas SR del estrés hídrico
prolongado.
El incremento gradual y continuo de las plantas CR pudo estar relacionado al
incremento de la edad fisiológica, unido al efecto del traslado de las
plantas al exterior de la casa de cultivo, este último relacionado con
la exposición durante tiempo prolongado a condiciones de alta intensidad
lumínica y a un déficit hídrico moderado, lo cual está
asociado a la disminución de los niveles de ácidos orgánicos
a los 45 días en este mismo grupo.
Las plantas CAM terrestres presentan un aumento del contenido de agua de tallos
y hojas en respuesta a condiciones limitantes de humedad, este incremento se
relaciona con la síntesis de tejidos de reserva de agua y con el aumento
del potencial osmótico de las células, este último provocado
por el influjo de ácido málico y carbohidratos que se almacenan
en el interior de las grandes y dominantes vacuolas (12).
El incremento del potencial osmótico explica la disminución del
contenido de agua en las plantas SR durante el periodo de déficit hídrico
(Figura 2A), el incremento
significativo de las plantas SR con el restablecimiento del riego y el aumento
gradual de las plantas CR durante el experimento, lo cual está relacionado
con un decremento o incremento de los niveles de ácidos orgánicos
y por ende de ácido málico según el caso; aunque esta conjetura
no es consistente con la disminución de los niveles de AO en las plantas
CR a los 45 días, de modo que con el establecimiento del riego hasta
la saturación del sustrato dicha disminución pudo estar asociada
a una menor magnitud de expresión CAM.
Efecto del déficit hídrico en la actividad de Superóxido
dismutasa
En la Figura 5 se observa un aumento de la actividad de SOD
en las plantas SR a los 15 días de evaluación sin diferencias
significativas entre ellas a los 30 días, a partir de este momento mostraron
una disminución en la actividad de SOD. El déficit hídrico
aplicado a las plantas SR provocó un aumento de la actividad SOD, esto
indica que durante los momentos de déficit hídrico (15 y 30 días)
las plantas SR debieron incrementar el grado de expresión ya que se ha
demostrado que la expresión de CAM está relacionado con rutas
de estrés oxidativo (4).
Con el restablecimiento del riego hasta condiciones de saturación del sustrato se observó una disminución significativa de la actividad de SOD en las plantas SR, esto implica una disminución de la magnitud de expresión CAM en este grupo. Las plantas CR no alcanzaron los valores de actividad SOD de las plantas SR en ninguno de los momentos de déficit hídrico, lo cual implica que las plantas SR presentaron una mayor capacidad para la eliminación del radical superóxido comparadas con las plantas CR, además indica que a los 15 días las plantas SR presentaron mejor preparación para atenuar el efecto negativo de las drásticas condiciones ambientales de campo comparado con las condiciones ambientales de aclimatización.
CONCLUSIONES
BIBLIOGRAFÍA
1.
Rodríguez, E. R.; López, D.; Aragón, C.; Becquer, R.; Pino,
Y.; Garza, Y.; Podestá, F. E.; González, O. J. L. y Rodríguez,
R. ‘‘CAM Metabolic Changes of ‘MD-2’ Pineapple Grown Under High and Low Light’’.
Pineapple News, vol. 21, 2014, pp. 27-34.
2. González-Olmedo, J. L.; Fundora, Z.; Molina, L. A.; Abdulnour, J.;
Desjardins, Y. y Escalona, M. ‘‘New contributions to propagation of pineapple
(Ananas comosus L. Merr) in temporary immersion bioreactors’’. In
Vitro Cellular & Developmental Biology - Plant, vol. 41, no. 1, enero
de 2005, pp. 87-90, ISSN 1054-5476, 1475-2689, DOI 10.1079/IVP2004603.
3. Aragón, C.; Carvalho, L.; González, J.; Escalona, M. y Amancio,
S. ‘‘The physiology of ex vitro pineapple (Ananas comosus
L. Merr. var MD-2) as CAM or C3 is regulated by the environmental conditions’’.
Plant Cell Reports, vol. 31, no. 4, 2 de diciembre de 2011, pp. 757-769,
ISSN 0721-7714, 1432-203X, DOI 10.1007/s00299-011-1195-7.
4. Aragón, C.; Pascual, P.; González, J.; Escalona, M.; Carvalho,
L. y Amancio, S. ‘‘The physiology of ex vitro pineapple (Ananas
comosus L. Merr. var MD-2) as CAM or C3 is regulated by the environmental
conditions: proteomic and transcriptomic profiles’’. Plant Cell Reports,
vol. 32, no. 11, 20 de agosto de 2013, pp. 1807-1818, ISSN 0721-7714, 1432-203X,
DOI 10.1007/s00299-013-1493-3.
5. Yanes, P. E.; González, O. J. y Sánchez, R. R. ‘‘A technology
of acclimatization of pineapple vitroplants’’. Pineapple News, vol.
15, 2000, pp. 24-35.
6. Villalobos, A.; González, J.; Santos, R. y Rodríguez, R. ‘‘Changes
in pineapple plantlets (Ananas comosus (L.) Merr.) during acclimatization’’.
Ciência e Agrotecnologia, vol. 36, no. 6, 2012, pp. 624-630,
ISSN 1981-1829.
7. Rodríguez, R.; Becquer, R.; Pino, Y.; Rodríguez-Escriba, R.
C. y López, D. ‘‘Introduction of pineapple vitroplantas to field conditions
in collaboration with farmers. Preliminary results’’. Pineapple News,
vol. 20, 2013, pp. 51-56.
8. Herrera, A. ‘‘Crassulacean acid metabolism and fitness under water deficit
stress: if not for carbon gain, what is facultative CAM good for?’’. Annals
of Botany, vol. 103, no. 4, 2 de enero de 2009, pp. 645-653, ISSN 0305-7364,
1095-8290, DOI 10.1093/aob/mcn145, PMID: 18708641.
9. Prigge, M. y Guriérrez-Soto, M. V. ‘‘Pineapple photosynthesis and
leaf sap pH as a surrogate of CAM performance in the field. A reseach advance’’.
Pineapple News, vol. 21, 2014, pp. 18-23.
10. Nievola, C. C.; Kraus, J. E.; Freschi, L.; Souza, B. M. y Mercier, H. ‘‘Temperature
determines the occurrence of CAM or C3 photosynthesis in pineapple plantlets
grown in vitro’’. In Vitro Cellular & Developmental Biology
- Plant, vol. 41, no. 6, noviembre de 2005, pp. 832-837, ISSN 1054-5476,
1475-2689, DOI 10.1079/IVP2005694.
11. Freschi, L.; Rodrigues, M. A.; Domingues, D. S.; Purgatto, E.; Sluys, M.-A.
V.; Magalhaes, J. R.; Kaiser, W. M. y Mercier, H. ‘‘Nitric Oxide Mediates the
Hormonal Control of Crassulacean Acid Metabolism Expression in Young Pineapple
Plants’’. Plant Physiology, vol. 152, no. 4, 1 de abril de 2010, pp.
1971-1985, ISSN 1532-2548, DOI 10.1104/pp.109.151613, PMID: 20147491.
12. Lüttge, U. ‘‘Ecophysiology of Crassulacean Acid Metabolism (CAM)’’.
Annals of Botany, vol. 93, no. 6, 6 de enero de 2004, pp. 629-652,
ISSN 0305-7364, 1095-8290, DOI 10.1093/aob/mch087, PMID: 15150072.
13. Escalona, M.; Lorenzo, J. C.; González, B.; Daquinta, M.; González,
J. L.; Desjardins, Y. y Borroto, C. G. ‘‘Pineapple (Ananas comosus
L. Merr) micropropagation in temporary immersion systems’’. Plant Cell Reports,
vol. 18, no. 9, mayo de 1999, pp. 743-748, ISSN 0721-7714, 1432-203X, DOI 10.1007/s002990050653.
14. Villalobo, A.; González, J.; Santos, R. y Rodríguez, R. ‘‘Morpho-physiological
changes in pineapple plantlets (Ananas comosus (L.) merr.) during acclimatization’’.
Ciência e Agrotecnologia, vol. 36, no. 6, diciembre de 2012,
pp. 624-630, ISSN 1413-7054, DOI 10.1590/S1413-70542012000600004.
15. Porra, R. J. ‘‘The chequered history of the development and use of simultaneous
equations for the accurate determination of chlorophylls a and b’’ [en línea].
En: eds. Govindjee, Beatty J. T., Gest H., y Allen J. F., Discoveries in
Photosynthesis, (ser. Advances in Photosynthesis and Respiration, no. ser.
20), edit. Springer Netherlands, 2005, pp. 633-640, ISBN 978-1-4020-3323-0,
DOI: 10.1007/1-4020-3324-9_56, [Consultado: 25 de enero de 2016], Disponible
en: <http://link.springer.com/chapter/10.1007/1-4020-3324-9_56>.
16. Rodríguez, E. R. C.; Rodríguez, R.; López, D.; Lorente,
G. Y.; Pino, Y.; Aragón, C. E.; Garza, Y.; Podestá, F. E. y González-Olmedo,
J. L. ‘‘High Light Intensity Increases the CAM Expression in «MD-2»
Micro-Propagated Pineapple Plants at the End of the Acclimatization Stage’’.
American Journal of Plant Sciences, vol. 06, no. 19, 2 de diciembre
de 2015, p. 3109, ISSN 2158-2750, DOI 10.4236/ajps.2015.619303.
17. Ceusters, J.; Borland, A. M.; Taybi, T.; Frans, M.; Godts, C. y Proft, M.
P. D. ‘‘Light quality modulates metabolic synchronization over the diel phases
of crassulacean acid metabolism’’. Journal of Experimental Botany,
vol. 65, no. 13, 7 de enero de 2014, pp. 3705-3714, ISSN 0022-0957, 1460-2431,
DOI 10.1093/jxb/eru185, PMID: 24803500.
18. Bradford, M. M. ‘‘A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding’’. Analytical
Biochemistry, vol. 72, no. 1, 7 de mayo de 1976, pp. 248-254, ISSN 0003-2697,
DOI 10.1016/0003-2697(76)90527-3.
19. StatPoint Technologies. Statgraphics Centurion [en línea].
(ser. Centurion), versión 16.1 (XV), [Windows], 21 de mayo de 2010, Disponible
en: <http://statgraphics-centurion.software.informer.com/download/>.
20. Borland, A. M. y Yang, X. ‘‘Informing the improvement and biodesign of crassulacean
acid metabolism via system dynamics modelling’’. New Phytologist, vol.
200, no. 4, 1 de diciembre de 2013, pp. 946-949, ISSN 1469-8137, DOI 10.1111/nph.12529.
21. Borland, A. M.; Wullschleger, S. D.; Weston, D. J.; Hartwell, J.; Tuskan,
G. A.; Yang, X. y Cushman, J. C. ‘‘Climate-resilient agroforestry: physiological
responses to climate change and engineering of crassulacean acid metabolism
(CAM) as a mitigation strategy’’. Plant, Cell & Environment, vol. 38,
no. 9, 1 de septiembre de 2015, pp. 1833-1849, ISSN 1365-3040, DOI 10.1111/pce.12479.
22. Borland, A. M.; Barrera Zambrano, V. A.; Ceusters, J. y Shorrock, K. ‘‘The
photosynthetic plasticity of crassulacean acid metabolism: an evolutionary innovation
for sustainable productivity in a changing world’’. New Phytologist,
vol. 191, no. 3, 1 de agosto de 2011, pp. 619-633, ISSN 1469-8137, DOI 10.1111/j.1469-8137.2011.03781.x.
23. Garcia, T. M.; Heyduk, K.; Kuzmick, E. y Mayer, J. A. ‘‘Crassulacean acid
metabolism biology’’. New Phytologist, vol. 204, no. 4, 1 de diciembre
de 2014, pp. 738-740, ISSN 1469-8137, DOI 10.1111/nph.13127.
24. Nelson, E. A. y Sage, R. F. ‘‘Functional constraints of CAM leaf anatomy:
tight cell packing is associated with increased CAM function across a gradient
of CAM expression’’. Journal of Experimental Botany, vol. 59, no. 7,
1 de mayo de 2008, pp. 1841-1850, ISSN 0022-0957, 1460-2431, DOI 10.1093/jxb/erm346,
PMID: 18256047.
25. Edwards, E. J. y Ogburn, R. M. ‘‘Angiosperm Responses to a Low-CO2
World: CAM and C4 Photosynthesis as Parallel Evolutionary Trajectories’’.
International Journal of Plant Sciences, vol. 173, no. 6, 2012, pp.
724-733, ISSN 1058-5893, DOI 10.1086/666098.
26. Herrera, A. ‘‘Crassulacean acid metabolism-cycling in Euphorbia milii’’.
AoB Plants, vol. 5, 1 de enero de 2013, p. plt014, ISSN 2041-2851,
DOI 10.1093/aobpla/plt014, PMID: 23596548.
27. Quezada, I. M. y Gianoli, E. ‘‘Crassulacean acid metabolism photosynthesis
in Bromeliaceae: an evolutionary key innovation’’. Biological Journal of
the Linnean Society, vol. 104, no. 2, 2011, pp. 480–486, ISSN 1095-8312.
28. Reyes, G. C.; Mejia, C. M. y Griffiths, H. ‘‘High but not dry: diverse epiphytic
bromeliad adaptations to exposure within a seasonally dry tropical forest community’’.
New Phytologist, vol. 193, no. 3, 1 de febrero de 2012, pp. 745-754,
ISSN 1469-8137, DOI 10.1111/j.1469-8137.2011.03946.x.
29. Matsuo, T.; Totoki, Y.; Suemitsu, H.; Matsuo, T.; Totoki, Y. y Suemitsu,
H. ‘‘Adaptive Estimation of Biological Rhythm in Crassulacean Acid Metabolism
with Critical Manifold, Adaptive Estimation of Biological Rhythm in Crassulacean
Acid Metabolism with Critical Manifold’’. ISRN Applied Mathematics,
vol. 2013, 18 de junio de 2013, p. 9, ISSN 2090-5564, DOI 10.1155/2013/856404.
30. Niewiadomska, E.; Bilger, W.; Gruca, M.; Mulisch, M.; Miszalski, Z. y Krupinska,
K. ‘‘CAM-related changes in chloroplastic metabolism of Mesembryanthemum
crystallinum L.’’. Planta, vol. 233, no. 2, 3 de noviembre de
2010, pp. 275-285, ISSN 0032-0935, 1432-2048, DOI 10.1007/s00425-010-1302-y.
31. Davies, B. N. y Griffiths, H. ‘‘Competing carboxylases: circadian and metabolic
regulation of Rubisco in C3 and CAM Mesembryanthemum crystallinum L.’’.
Plant, Cell & Environment, vol. 35, no. 7, 1 de julio de 2012,
pp. 1211-1220, ISSN 1365-3040, DOI 10.1111/j.1365-3040.2012.02483.x.
32. Bartholomew, D. P.; Paull, R. E. y Rohrbach, K. G. The Pineapple: Botany,
Production, and Uses. edit. CABI, 20 de noviembre de 2002, 336 p., ISBN
978-0-85199-979-1.
33. Borland, A. M.; Hartwell, J.; Weston, D. J.; Schlauch, K. A.; Tschaplinski,
T. J.; Tuskan, G. A.; Yang, X. y Cushman, J. C. ‘‘Engineering crassulacean acid
metabolism to improve water-use efficiency’’. Trends in Plant Science,
vol. 19, no. 5, 1 de mayo de 2014, pp. 327-338, ISSN 1360-1385, DOI 10.1016/j.tplants.2014.01.006,
PMID: 24559590.
Recibido: 15 de
mayo de 2015
Aceptado: 18 de enero de 2016
René C. Rodríguez Escriba, Centro de Bioplantas. Universidad de Ciego de Ávila. Ciego de Ávila, Cuba. Email: renecarlos@bioplantas.cu