
http://dx.doi.org/10.13140/RG.2.1.1642.2642
Efecto del Pectimorf® en el enraizamiento y la aclimatización in vitro de brotes de papaya (Carica papaya L.) cultivar Maradol Roja
Efect of Pectimorf® on rooting and in vitro acclimatization of papaya (Carica papaya L.) shoots cultivar Maradol Roja
M.Cs. Laisyn Posada-Pérez,I Yenny Padrón-Montesinos,I Dr.C. Justo González-Olmedo,II Dr.C. Romelio Rodríguez-Sánchez,II Dr.C, Raul Barbón-Rodriguez,I M.Cs. Osvaldo Norman-Montenegro,III Rene C. Rodríguez-Escriba,II Dr.C. Rafael Gómez-KoskyI
IInstituto de Biotecnología de las Plantas, Universidad Central “Marta Abreu” de Las Villas, arretera a Camajuaní, km 5,5, Santa Clara, Villa Clara, Cuba.
IICentro de Bioplantas, Universidad de Ciego de Ávila, carretera a Morón, km 9, Ciego de Ávila, Cuba.
IIICentro de Bioactivos Químicos, Universidad Central “Marta Abreu” de Las Villas, carretera a Camajuaní, km 5,5, Santa Clara, Villa Clara, Cuba.
RESUMEN
La propagación in vitro de la papaya ha sido exitosamente informada por diferentes investigadores; sin embargo, los problemas en los procesos de enraizamiento y aclimatización aún no han sido completamente resueltos. Los bajos porcentajes de enraizamiento y la baja supervivencia de las plantas en condiciones de aclimatización continúan siendo un problema para el desarrollo de un eficiente protocolo de propagación in vitro en este cultivo. El empleo del regulador del crecimiento cubano Pectimorf® podría ser utilizado para minimizar este problema. En el trabajo se realizaron dos experimentos que tuvieron como tratamientos dos concentraciones de sacarosa (0 y 10 g L-1), dos de la auxina AIB (0 y 2 mg L-1) y cinco concentraciones de Pectimorf® (3, 5, 7, 9 y 12 mg L-1). Se emplearon como controles: el medio de cultivo de enraizamiento compuesto por las sales MS al 50 %; 2 mg L-1 de AIB; 0,4 mg L-1 de tiamina; 40 g L-1 de sacarosa; 7 g L-1 de agar y el mismo medio de cultivo sin sacarosa, sin Pectimorf® pero con zeolita como soporte. Los resultados demostraron que el Pectimorf® tuvo un efecto positivo en el enraizamiento y la aclimatización in vitro de los brotes de papaya. La acción sinérgica del AIB con 9 mg L-1 de Pectimorf® permitió obtener plantas in vitro con mayor área foliar, masa fresca, número de raíces, tasa fotosintética y conductancia estomática; lo cual unido a un alto porcentaje de enraizamiento y un menor porcentaje de estomas abiertos permitió alcanzar un 76,2 % de supervivencia en condiciones ex vitro.
Palabras clave: auxina, oligosacáridos, papaya, raíces, zeolita.
ABSTRACT
In vitro propagation of papaya has been successfully reported by different researchers, however the problems in the process of rooting and acclimatization have not been completely resolved yet. The low percentage of rooting and low survival of plants under conditions of acclimatization remain a problem for the development of an efficient micropropagation protocol for this crop. Employment growth regulator cuban Pectimorf® could be used to minimize this problem. In two experiments that were conducted as treatments are made up of two concentrations of sucrose (0 and 10 g L-1), two of auxin AIB (0 and 2 mg L-1) and five Pectimorf® concentrations (3, 5, 7, 9 and 12 mg L-1). Were used as controls: the culture medium rooting composed of MS salts 50 %, 2 mg L-1 IBA, 0,4 mg L-1 thiamine, sucrose 40 g L-1 agar 7 g L-1 and the same culture medium without sucrose and Pectimorf®, but with zeolite as carrier. The results showed that the Pectimorf® had a positive effect on rooting and in vitro acclimatization of papaya shoot. The synergistic action of AIB with 9,0 mg L-1 Pectimorf® allowed to obtain in vitro plants with greater leaf area, fresh weight, number of roots, photosynthetic rate and stomatal conductance, which together with a high percentage of rooting and less percentage of open stomata allowed to reach a 76,2 % survival ex vitro conditions.
Key words: auxin, oligosaccharides, papaya, roots, zeolite.
INTRODUCCIÓN
La papaya (Carica papaya L.) es originaria de América Central y se caracteriza por ser un cultivo que en corto tiempo y de forma continua durante todo un año, es productivo desde el punto de vista económico. Su alto rendimiento y valor nutritivo, la sitúan entre las joyas de los frutos tropicales con importantes aplicaciones alimenticias y medicinales (1).
El cultivar más importante de papaya en Cuba sigue siendo la Maradol Roja, con un área total cultivada de 5 396 ha y una producción de 105 562 toneladas en el año 2015A.
Aunque la regeneración de plantas de papaya vía embriogénesis somática ha sido exitosa, el mayor problema que existe a nivel mundial en el cultivo in vitro es la aclimatización de las plantas regeneradas (2).
Los procesos de propagación in vitro se implementan en grandes espacios físicos para el cultivo de las plantas; fundamentalmente en estantes que contienen frascos de cultivo cerrados con medios de cultivo nutritivos en su interior, para el crecimiento de las plantas y en condiciones de asepsia, para evitar las contaminaciones por hongos o bacterias oportunistas. Todas estas condiciones hacen que las plantas cultivadas in vitro sean muy sensibles a cambios ambientales abruptos, como los que ocurren en el paso a las condiciones ex vitro en ambientes naturales (3).
Como resultado del ambiente in vitro, las plantas presentan una anatomía y fisiología diferente a las que son cultivadas en condiciones de campo o casas de cultivo (4–6). Los desórdenes observados afectan todos los órganos de la planta, aunque no todos tienen el mismo peso sobre el comportamiento ex vitro. Dentro de estos desórdenes se encuentran el pobre desarrollo del aparato fotosintético de la cutícula de las hojas, la emisión de raíces no funcionales sin conexión con los haces conductores y otros más que pueden afectar la supervivencia de las plantas en la fase de aclimatización (7).
El término aclimatización es definido como la adaptación ambiental de las plantas obtenidas por cultivo de tejidos o propagación in vitro que han sido movidas a un nuevo ambiente, invernaderos o campo. Durante la aclimatización, el ambiente a las plantas le es cambiado gradualmente en el tiempo, comenzando con el cercano ambiente in vitro y terminando con el cercano ambiente en el invernadero o campo. La aclimatización realizada en el invernadero o campo bajo condiciones de sombra es llamada “aclimatización ex vitro”. En la propagación in vitro foto-autotrófica, la aclimatización puede ser también completada en el frasco de cultivo, lo cual es conocido como “aclimatización in vitro” (8).
Varios trabajos realizados hasta la fecha están encaminados a sistemas autotróficos y con ventilación forzada para preparar mejor a las plantas para su salida al exterior. El aumento de la intensidad de luz en los frascos, la concentración de CO2 y la ventilación forzada son aspectos que ayudan a favorecer una nutrición autotrófica, siempre eliminando la sacarosa, lo cual ayuda a disminuir los costos de producción en la propagación in vitro convencional; así como a reducir la contaminación microbiana en el medio de cultivo (8–10). Comúnmente cuando se elevan las concentraciones de CO2 se provoca un incremento de la fotosíntesis y el crecimiento vegetativo de las plantas en condiciones ex vitro (11).
Sin embargo, todos los estudios realizados con anterioridad para la propagación in vitro de la papaya revelan que los principales inconvenientes para propagar esta especie son el enraizamiento y la aclimatización de las plantas en invernadero (12, 13). La supervivencia puede alcanzar valores entre 65-70 % en estas condiciones durante los primeros siete días de cultivo ex vitro. Lo fundamental es que las plantas formen un buen sistema radical, debido a que su nutrición en gran parte dependerá de la funcionalidad de sus raíces. Es por esto que se deben establecer nuevas estrategias biotecnológicas para aumentar la eficiencia de los protocolos de propagación in vitro de plántulas de papaya, cultivar Maradol Roja.
La introducción de sustancias activas de producción nacional en la metodología de regeneración in vitro de plantas de papaya, pudiera constituir una alternativa para mejorar el enraizamiento in vitro. Dentro de estas sustancias activas de producción nacional se puede mencionar el Pectimorf® “mezcla de (1-4) α-D-oligogalacturónidos, con grado de polimerización de entre 9 y 16” (14).
El Pectimorf® se reconoce como un nuevo biorregulador cubano, obtenido a partir de residuos de la industria citrícola, cuyo principio activo es una mezcla de oligosacáridos de origen péptico. La capacidad del Pectimorf® para inducir y desarrollar el enraizamiento e incrementar de forma notable el desarrollo y vigor de las plantas in vitro de los diferentes cultivos, lo validan como una alternativa promisoria en la biotecnología vegetal (14–16).
Por todo lo anteriormente planteado el objetivo del presente trabajo fue determinar el efecto del producto cubano Pectimorf® en el enraizamiento y la aclimatización in vitro de plantas de papaya, bioregulador no utilizado hasta el momento para este fin en esta especie.
MATERIALES Y MÉTODOS
Las investigaciones se realizaron en el Instituto de Biotecnología de las Plantas, de la Universidad Central “Marta Abreu” de Las Villas, en Santa Clara, Cuba.
Como material vegetal se emplearon brotes in vitro de papaya (Carica papaya L.) del cultivar Maradol Roja regenerados a partir de embriones somáticos. Estos tenían cuatro subcultivo en el medio de cultivo de elongación (17), que contenía las sales: MS al 100 % de su concentración; 1,0 mg L-1 de tiamina; 1,2 µM de 6 bencilamino purina (6 BAP); 1,5 µM de ácido naftalenacético (ANA); 100 mg L-1 de mioinositol; 30 g L-1 de sacarosa; 0,06 mg L-1 de vitamina B2 y 5 g L-1 de Agargel (Sigma Co) (18). Los brotes para los ensayos fueron seleccionados con una longitud entre 3,0–4,0 cm, a los cuales se les eliminaron las hojas basales y se le dejaron las cuatro nuevas últimas hojas.
Condiciones de cultivo
Para ambos experimentos se emplearon frascos de cultivo de vidrio de 250 mL de capacidad, con tapa plástica transparente en el tratamiento control con agar. Los frascos con el medio de cultivo líquido y zeolita fueron cubiertos con una lámina de papel de aluminio. A todos se les añadió 30 mL de medio de cultivo. Cada tratamiento tuvo 33 repeticiones, colocando dos brotes por frasco de cultivo que fueron cultivados en cuartos climatizados a una temperatura de 27±2 °C, con luz solar con un fotoperíodo de 13/11 h de luz/oscuridad y con un rango de flujo de fotones fotosintéticos entre 48,0 y 62,5 µmol m-2 s-1; medido con un Luxómetro Extech 401025 (Extech Instruments, EUA). Los experimentos fueron repetidos dos veces.
A los tres días de cultivo, se comenzó a abrirle un orificio a la lámina de papel de aluminio que cubrían los frascos de cultivo en los distintos tratamientos y experimentos, con el objetivo de incrementar la ventilación, con el auxilio de una pinza estéril. El segundo orificio se abrió a los cuatro días de haber abierto el primero (Figura 1).
Efecto de la combinación sacarosa, el AIB y el Pectimorf® en el enraizamiento y la aclimatización in vitro
El objetivo de este primer experimento fue determinar el efecto de la combinación de la sacarosa y la presencia o no del regulador del crecimiento ácido indol butírico (AIB) y varias concentraciones de Pectimorf®, para el enraizamiento de los brotes in vitro. Se utilizó como soporte el mineral zeolita estéril, el alumino-silicato natural con excelentes propiedades de intercambio iónico, con un alto poder de absorción y granulación 1-3 mm (Tabla I). Se adicionó a cada frasco de cultivo de vidrio 97 g de este mineral y fueron tapados con una lámina de papel de aluminio (Figura 1).

Se estudiaron tres concentraciones de Pectimorf® (3, 5, 7 mg L-1) con y sin presencia de sacarosa (0 y 10 g L-1) y AIB (2 mg L-1). Se emplearon como tratamientos controles: el medio de cultivo de enraizamiento propuesto anteriormente (17), que estaba compuesto por las sales MS al 50 % de su concentración; 2 mg L-1 de AIB; 0,4 mg L-1 de tiamina; 40 g L-1 de sacarosa; 7 g L-1 de agar, pH 5,8 previo al autoclaveado y el mismo medio de cultivo pero con zeolita; 2 mg L-1 de AIB y sin sacarosa.
Efecto de la combinación AIB-Pectimorf® sin sacarosa en el enraizamiento y aclimatización in vitro
El objetivo de este segundo experimento fue evaluar concentraciones superiores de Pectimorf® en combinación con el AIB, tomando en cuenta los resultados del experimento anterior. Se emplearon las siguientes concentraciones 7, 9 y 12 mg L-1 de Pectimorf®. En este experimento se empleó como tratamiento control el medio de cultivo de enraizamiento descrito anteriormente suplementado solamente con 2 mg L-1 de AIB, y zeolita como soporte.
En este experimento un grupo de 20 plantas por cada tratamiento fueron llevadas a condiciones ex vitro en la fase de aclimatización para evaluar la supervivencia.
Para las evaluaciones de indicadores morfológicos y fisiológicos de las plantas se seleccionaron al azar 20 brotes por tratamiento a partir de los 37 días de cultivo. Además, se evaluó el porcentaje de contaminación en el 100 % de los frascos de cultivo con los dos oficios y los controles.
Evaluaciones de variables morfológicas y fisiológicas
Al terminar los experimentos in vitro se le realizaron las siguientes evaluaciones morfológicas a los brotes y a las plantas: altura de la planta (cm); número de hojas; número de entrenudos; enraizamiento (%); número de raíces; longitud de la raíz (cm); masa fresca de la planta in vitro (gMF); presencia o no de callo basal.
Además, para el segundo experimento se evaluó el número de estomas por mm2 y porcentaje de estomas abiertos y cerrados, tomado de las muestras a las 12 del mediodía a diez plantas in vitro (37 días de cultivo) y diez ex vitro (siete días en fase de aclimatización), el área foliar (por el método propuesto para plantas de papaya) (19), unido a los indicadores fisiológicos como la actividad fotosintética neta, la transpiración total y la conductancia estomática.
Observación de los estomas
La visualización de los estomas se realizó en la parte abaxial de la hoja. Se utilizó el método de la réplica modificado por Engleman (20); que consiste en poner una gota de pegamento instantáneo Kola Loka® (Kola Loka SA de CV, México) en un portaobjetos, posteriormente presionar la hoja por un minuto sobre la gota y enseguida despegar del portaobjetos. Las impresiones se llevaron al laboratorio donde se observaron en un microscopio óptico ANJUE N-800 (Shanghai, China) que tenía adaptada una cámara digital HDCE-50B (Alltion, China). Se fotografiaron tres campos elegidos al azar por tratamiento para el conteo de los estomas.
Determinación de la fotosíntesis, transpiración total y conductancia estomática
Para las determinaciones se utilizaron hojas totalmente expandidas en la misma posición (hojas dos y tres) en los brotes y plantas in vitro al final del experimento, entre cuatro y cinco horas después del principio del fotoperíodo. Se realizaron las determinaciones por tratamiento en 12 plantas con diez mediciones en cada una para un total de 120 mediciones. La capacidad fotosintética máxima (µmol CO2 m-2 s-1), la transpiración total (mmol H2O m-2 s-1) y la conductancia estomática (mmol m-2 s-1) se midieron con el equipo CIRAS-2 (Sistema Portátil de Fotosíntesis, Reino Unido) acoplado a una cubeta universal PLC6 2,5 cm2. El área de la cubeta se cubrió completamente con la hoja (1,7 cm2). La concentración del dióxido de carbono, la temperatura del aire y la humedad relativa (80-90 %) fueron valores ambientales. Para el trabajo con el equipo la luz fue fijada a una intensidad de 900 µmol m-2 s-1. Las mediciones fueron siempre realizadas a todas las plantas in vitro entre las 9:00 a 10:00 a.m.
Además se calculó la eficiencia en el uso del agua de la fotosíntesis mediante la fórmula: asimilación neta de CO2 / conductancia estomática (21).
Condiciones de aclimatización ex vitro
Las plantas para su crecimiento se cultivaron bajo un fotoperíodo de 12 h de luz/12 h de oscuridad, con una temperatura media durante el día de 30±2 ºC y una humedad relativa de 70-75 %. La intensidad lumínica osciló entre 224 y 457 µmol m-2 s-1, la cual se midió con un Luxómetro Extech 401025 (Extech Instruments, EUA).
Se utilizó un sustrato compuesto por zeolita (Tabla I) y compost de cachaza de caña de azúcar (Saccharum spp, híbrido) (9:1, v/v). Estos componentes fueron colocados en macetas de plástico de 500 mL de volumen total. Primero se depositó en el fondo el compost y encima la zeolita para garantizar una buena aireación de las raíces de las plantas de papaya. El riego fue realizado de forma manual por aspersión dos veces al día. Las plantas fueron cubiertas por un frasco de vidrio transparente durante cinco días para garantizar una alta humedad relativa mayor del 90 % y un sombreo del 70 % con una malla sarán negra.
Se utilizaron por tratamiento 20 plantas con la combinación AIB con 9 y 12 mg L-1 de Pectimorf® y los controles sin Pectimorf® en zeolita y agar. El porcentaje de supervivencia (%) se determinó contando las plantas que se mantenían vivas en el momento de evaluación (siete días) (22).
Análisis estadístico
Para los análisis estadísticos se utilizó el paquete de programas SPSS para Windows versión 21 (23). Para el análisis de la normalidad de las variables se utilizó el test de Shapiro Wilk, para la comparación entre las medias se aplicó la alternativa no paramétrica del Análisis de Varianza, el test de Kruskal-Wallis y para la comparación entre parejas de grupos se utilizó el test no paramétrico de Mann-Whitney.
RESULTADOS Y DISCUSIÓN
Efecto de la combinación sacarosa, el AIB y el Pectimorf® en el enraizamiento y la aclimatización in vitro
El tratamiento control con zeolita (sin sacarosa y 2 mg L-1 de AIB) tuvo una respuesta superior en el porcentaje de enraizamiento respecto al resto de los tratamientos. En las demás variables morfológicas evaluadas tuvo valores iguales y superiores con diferencias significativas, en varios de los tratamientos. Ninguna de las concentraciones de Pectimorf® empleadas en este primer experimento (3, 5 y 7 mg L-1) superaron los resultados del tratamiento control zeolita; sin embargo, es de destacar que todos fueron superiores al control con agar con 40 g L-1 de sacarosa (Tabla II).
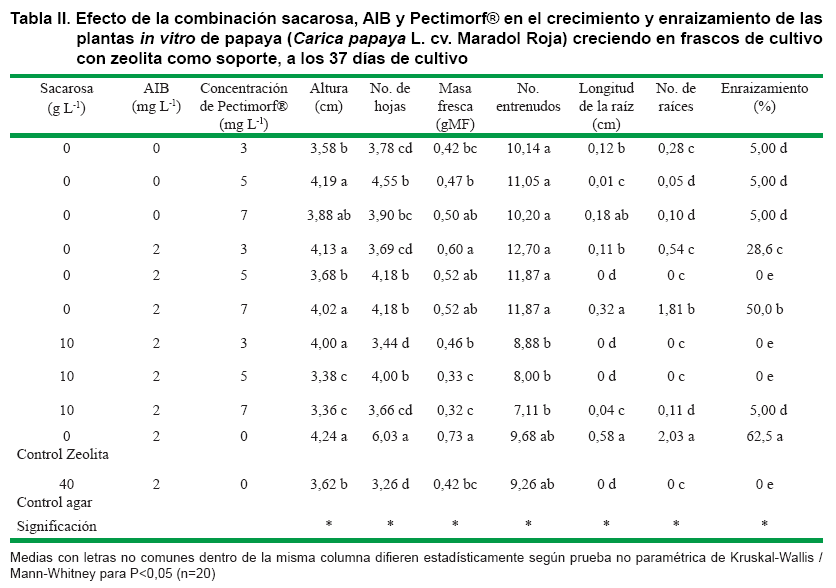
Para las variables relacionadas con el enraizamiento tales como, número de raíces y porcentaje de enraizamiento, los valores alcanzados por el tratamiento control zeolita con 2 mg L-1 de AIB, fueron superiores al resto de los tratamientos con diferencias significativas.
En este experimento las concentraciones de Pectimorf® estudiadas en combinación o no con la auxina AIB, no estimularon la longitud ni el número de la raíz. Solo se alcanzó un 50,0 % de enraizamiento en el tratamiento sin sacarosa, 2 mg L-1 de AIB y 7 mg L-1 de Pectimorf® con valor inferior al 62,5 % del tratamiento control con zeolita con 2 mg L-1 de AIB (Tabla II).
En los tratamientos donde no estuvo presente la sacarosa en el medio de cultivo, las plantas crecieron y se desarrollaron de forma normal comparados con los tratamientos con sacarosa (10 g L-1 y el control 40 g L-1). Esto supone que las plantas in vitro de papaya en estas condiciones de cultivo tuvieron un comportamiento fotoautotrófico (Tabla II). Sin embargo, es importante destacar que con la presencia de la sacarosa en el medio de cultivo y a pesar de la presencia de ambos estimuladores del crecimiento el porcentaje de enraizamiento fue muy bajo (5,0 %) o nulo.
No siempre en todas las plantas cultivadas in vitro la eliminación de la sacarosa en el medio de cultivo presupone el desarrollo de un buen sistema de raíces.
Se ha informado que plantas creciendo en medio de cultivo con sacarosa mostraron un mayor número y raíces más largas que en el tratamiento sin sacarosa, en la orquídea del género Doritaenopsis (24).
En el cultivo in vitro de la papaya es conocido el uso del regulador del crecimiento AIB para la formación de raíces (13, 17, 25); sin embargo, este regulador del crecimiento en el medio de cultivo semisólido estimula la formación de un callo basal que impide o hace infuncional algunas de las raíces que se forman, por no tener conexión con el tallo. En ninguno de los tratamientos con zeolita como soporte se formó callo basal en los brotes, a pesar de estar presente el AIB a una concentración de 2 mg L-1 y en combinación con el Pectimorf®. En el cultivo in vitro de la papaya solo ha sido estudiado el uso de Pectimorf® a baja concentración (1 mg L-1) en la propagación in vitro (organogénesis) en combinación con el 6 BAP en las fases de establecimiento y multiplicaciónB.
La zeolita también permitió un mejor crecimiento y desarrollo de las raíces, dado por la mayor aireación que reciben durante su crecimiento in vitro. Resultados estos que anteriormente no se habían obtenido para este cultivo. No obstante en el cultivo de la papaya in vitro se ha utilizado la vermiculita como soporte para el enraizamiento de los brotes.
El uso de la vermiculita combinado con el AIB para el enraizamiento de distintas variedades de papaya ha sido informado por varios autores (25, 26). Los mismos señalan el efecto beneficioso de la vermiculita para lograr altos porcentajes de enraizamiento (80-90 %). Sin embargo, algunos autores señalan el efecto superior del AIB sobre otras auxinas (ANA; AIA) para lograr el enraizamiento de brotes in vitro de papaya, pero en medio de cultivo semisólido (13).
Respecto a la contaminación microbiana, los niveles alcanzados fueron de 10,0-15,0 % de los frascos de cultivo, donde el medio tenía sacarosa, a los 37 días de cultivo. En el resto de los tratamientos sin la presencia de la sacarosa en el medio de cultivo los valores de contaminación fueron menores del 4,0 %.
Los resultados alcanzados en este primer experimento llevaron al estudio de concentraciones más altas de Pectimorf® para incrementar el porcentaje de enraizamiento de los brotes in vitro de papaya cv. Maradol Roja en presencia de AIB, sin sacarosa en el medio de cultivo y utilizando la zeolita como soporte.
Efecto de la combinación AIB-Pectimorf® sin sacarosa en el enraizamiento y aclimatización in vitro
Los tratamientos con las mayores concentraciones de Pectimorf® 9 y 12 mg L-1 (Figura 2), tuvieron una respuesta superior para la variable morfológica (área foliar) con diferencias significativas con el resto de los tratamientos. Respecto a la variable longitud de la raíz más larga, los valores más altos se alcanzaron en ambos tratamientos anteriormente señalados, pero sin diferencias estadísticas. No obstante, el mayor porcentaje de plantas con raíces (84,2 %) fue alcanzado con la concentración de 9 mg L-1 de Pectimorf® en complemento con la auxina AIB, siendo superior al resto de los tratamientos y al control, a los 37 días de cultivo (Tabla III).

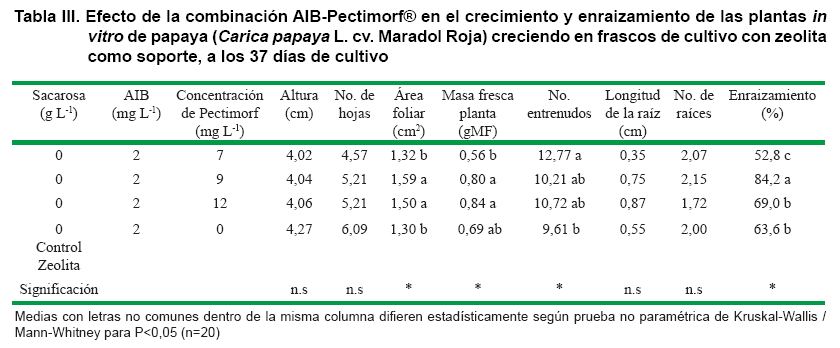
En el presente trabajo la mejor concentración de Pectimorf® que estimuló la formación de raíces en brotes de papaya coincide con iguales concentraciones utilizadas en otros cultivos empleando este bioproducto. Por ejemplo, en el cultivo de la yuca (Manihot esculenta L.)C, obtuvieron los mejores resultados con 9 mg L-1 de Pectimorf® para aumentar el enraizamiento de los brotes in vitro y su aclimatización. En el Spathiphyllium sp (15) lograron el estimuló del desarrollo radical de los brotes in vitro. También se observó la promoción de raíces secundarias en los pecíolos de la violeta (Saintpaulia ionantha), o sea, un efecto auxínico al emplear la mezcla de oligogalacturónicos a 10 mg L-1 (16) en comparación con la auxina ácido indol acético (AIA); y no incrementó el número de raíces, ni su longitud igual que ocurrió con los brotes in vitro de papaya.
Otros autores señalan el criterio de que el Pectimorf® ejerce una acción similar al de las auxinas o de una sinergia con la auxina presente en el medio de cultivo (27). Los resultados obtenidos en el presente trabajo demuestran la sinergia del Pectimorf® con la auxina AIB.
También en tabaco (Nicotiana tabacum L.) y Arabidopsis thaliana (L.) Heynh (28), utilizando 10 mg L-1 de Pectimorf® en combinación con la auxina AIB, pero en concentración más baja (0,5 mg L-1) a la del presente trabajo, alcanzaron los mejores resultados en el incremento de la longitud de la raíz. Estos autores informan que el Pectimorf® estimuló la elongación de la raíz primaria y desaceleró la formación de raíces laterales. El Pectimorf® a las concentraciones estudiadas en combinación con el AIB no estimuló la formación de un mayor número de raíces por brote in vitro en el cultivo de papaya (Tablas II y III).
Al analizar los resultados de las variables fisiológicas evaluadas (fotosíntesis, transpiración y conductancia estomática) los tratamientos con Pectimorf® tuvieron un efecto negativo sobre la actividad fotosintética de las plantas in vitro de papaya a los 37 días de cultivo.
El tratamiento control (zeolita y 2 mg L-1 de AIB) tuvo el mayor valor de fotosíntesis y conductancia estomática con diferencias significativas con el resto de los tratamientos; sin embargo, tuvo una mayor transpiración lo cual es negativo para la aclimatización de las plantas in vitro a las condiciones ex vitro ya que estas sufrirán una mayor pérdida de agua (Tabla IV).

Al analizar la eficiencia en el uso del agua de la fotosíntesis la concentración de 12 mg L-1 de Pectimorf® alcanzó el valor más alto, respecto a los demás tratamientos, lo que demuestra que las plantas de papaya estaban captando más CO2 con el concurso de menos agua (Tabla IV).
El tratamiento con 9 mg L-1 de Pectimorf® tuvo menor nivel de fotosíntesis y conductancia estomática respecto al control zeolita; pero superior al resto de las concentraciones de Pectimorf® con diferencias significativas, además de una menor transpiración que el control, teniendo una mejor respuesta en las condiciones ex vitro con altos valores de supervivencia para este cultivo (Tabla V).

Respecto a la relación estomas abiertos y cerrados así como el número total de estos por mm-2; los mayores porcentajes de estomas abiertos se alcanzaron en las plantas in vitro de los controles (soporte con zeolita y agar) con 35,0 % y 72,41 % respectivamente. Sin embargo, en los tratamientos con el Pectimorf® y la auxina AIB, los valores fueron inferiores, lo cual es un aspecto positivo para la adaptación posterior a condiciones ex vitro.
Esto permite afirmar que las plantas de papaya han tenido un proceso de aclimatización in vitro con el uso del Pectimorf®. También con la concentración de 9 mg L-1 de Pectimorf® el número de estomas por área fue inferior al resto de los tratamientos (Tabla V). Todo lo anterior, unido a un mayor porcentaje de plantas con raíces en el tratamiento con 9 mg L-1 de Pectimorf® permitió obtener a los siete días en condiciones de aclimatización ex vitro el mayor porcentaje de supervivencia (76,2 %); a pesar de tener el tratamiento con 12 mg L-1 una mayor eficiencia en el uso del agua de la fotosíntesis. Todo esto demuestra la importancia de obtener un alto número de plantas in vitro de papaya, con raíces para garantizar altos porcentajes de supervivencia ex vitro.
Es importante señalar la respuesta de las plantas del tratamiento control agar, las cuales no tenían raíces producto de las condiciones del cultivo in vitro con alta humedad relativa dentro del frasco de cultivo; estas tuvieron un alto porcentaje de estomas abiertos 72,4 % y a pesar de no tener un alto número de estomas por área, el 100 % de las plantas no sobrevivieron a las condiciones ex vitro de aclimatización (Tabla V).
Autores informan que al comparar la densidad estomática en hojas de plantas cultivadas in vitro con otras en condiciones de invernadero encontraron que la densidad estomática es más del doble en las plantas in vitro que las de invernadero (29).
Además, encontraron diferencias en el tamaño y el número de estomas. Las plantas in vitro de la especie leñosa Castanea sativa tuvieron una mayor proporción de estomas abiertos que las plantas de invernadero. En esta investigación fue posible cambiar esta proporción, debido a las condiciones empleadas que permitieron su aclimatización in vitro como fotoautotrófismo (medio de cultivo sin sacarosa), zeolita como soporte, la apertura en la cubierta de los frascos permitiendo una mayor aireación y alta supervivencia ex vitro de las plantas.
El Pectimorf® ha sido utilizado por varios autores para el enraizamiento en diferentes especies de plantas cultivadas in vitro (15, 16, 30). Todos estos autores señalan que la concentración adecuada del Pectimorf® como regulador del crecimiento para el cultivo in vitro de plantas es alrededor de los 10 mg L-1, lo cual apoya los resultados de este trabajo.
Este oligogalacturónido ejerció una acción de sinergia con la auxina, presente en el medio de cultivo. Lo que se demuestra en el enraizamiento de los brotes in vitro de papaya al alcanzar un 20,6 % más de brotes con raíces con la mejor combinación de AIB y Pectimorf® (9 mg L-1) respecto al control sin Pectimorf®. Parte de este efecto positivo en el crecimiento son atribuidos a los aumentos en las tasas de división celular. Sin embargo, los mecanismos que explican la estimulación del Pectimorf® en la división celular en las plantas superiores aún son desconocidos (28).
La acción sinérgica del AIB con 9 mg L-1 de Pectimorf®, unido a las condiciones de cultivo que se utilizaron, permitió obtener plantas in vitro de papaya con una mayor área foliar, masa fresca, mayor valor en el número de raíces, una elevada tasa fotosintética y conductancia estomática; lo cual unido a un alto porcentaje de enraizamiento (84,2 %) y un menor porcentaje de estomas abiertos con respecto al resto de los tratamientos permitió alcanzar un 76,2 % de supervivencia en condiciones ex vitro.
CONCLUSIONES
Notas al pie
AMINAG (2016) Estadísticas oficiales del Ministerio de la Agricultura. La Habana, Cuba. Cierre diciembre 2015.
BRoque, L. A. Y. Propagación in vitro de la papaya (Carica papaya L.) cv Maradol Roja: Una alternativa de solución para los productores habaneros. [Tesis de Doctorado], Universidad Agraria de La Habana, Facultad de Agronomía, La Habana, Cuba, 2004, 100 p.
CHernández, H. M.; Suarez, L.; Valcárcel, M. y López, M. ‘‘Empleo de una mezcla de oligogalacturónidos (Pectimorf) en la micropropagación de yuca (Manihot esculenta, Crantz) y Malanga (Colocasia sp.)’’. En: II Simposio Internacional de Raíces, Rizomas, Tubérculos, Plátanos, Bananos y Papaya, edit. Instituto Nacional de Investigaciones en Viandas Tropicales (INIVIT), Villa Clara, Cuba, 2013.
BIBLIOGRAFÍA
1. Zhang, J.; Shen, W.; Yan, P.; Li, X. y Zhou, P. ‘‘Factors that influence the yield and viability of protoplasts from Carica papaya L’’. African Journal of Biotechnology, vol. 10, no. 26, 2011, p. 5143, ISSN 1684–5315, DOI 10.5897/AJB10.1156.
2. Das, S. C.; Dinesh, M. R.; Das, A. y Suresh, C. P. ‘‘Studies on papaya cultivation in hilly tripura: Improved socio-economical status and their problems and prospects’’. Acta Horticulturae, no. 1022, marzo de 2014, pp. 165-173, ISSN 0567-7572, 2406-6168, DOI 10.17660/ActaHortic.2014.1022.21.
3. Chandra, S.; Bandopadhyay, R.; Kumar, V. y Chandra, R. ‘‘Acclimatization of tissue cultured plantlets: from laboratory to land’’. Biotechnology Letters, vol. 32, no. 9, 9 de mayo de 2010, pp. 1199-1205, ISSN 0141-5492, 1573-6776, DOI 10.1007/s10529-010-0290-0.
4. Hazarika, B. N. y Bora, A. ‘‘Hyperhydricity-A bottleneck to micropropagation of plants’’. Acta Horticulturae, no. 865, junio de 2010, pp. 95-101, ISSN 0567-7572, 2406-6168, DOI 10.17660/ActaHortic.2010.865.11.
5. Jausoro, V.; Llorente, B. E. y Apóstolo, N. M. ‘‘Structural differences between hyperhydric and normal in vitro shoots of Handroanthus impetiginosus (Mart. ex DC) Mattos (Bignoniaceae)’’. Plant Cell, Tissue and Organ Culture (PCTOC), vol. 101, no. 2, 13 de febrero de 2010, pp. 183-191, ISSN 0167-6857, 1573-5044, DOI 10.1007/s11240-010-9675-y.
6. Gutiérrez, M. A.; González, G. A. G.; Ascencio, C. A.; Rodríguez, G. B. y Li, W. L. Plant Somatic Embryogenesis: Some Useful Considerations. edit. INTECH Open Access Publisher, 2012, ISBN 978-953-51-0466-7.
7. Alvarez, C.; Sáez, P.; Sáez, K.; Sánchez-Olate, M. y Ríos, D. ‘‘Effects of light and ventilation on physiological parameters during in vitro acclimatization of Gevuina avellana mol’’. Plant Cell, Tissue and Organ Culture (PCTOC), vol. 110, no. 1, 22 de febrero de 2012, pp. 93-101, ISSN 0167-6857, 1573-5044, DOI 10.1007/s11240-012-0133-x.
8. Kozai, T. ‘‘Photoautotrophic micropropagation—environmental control for promoting photosynthesis’’. Prop Ornam Plants, vol. 10, 2010, pp. 188–204, ISSN 1311-9109.
9. Aragón, C.; Carvalho, L.; González, J.; Escalona, M. y Amancio, S. ‘‘The physiology of ex vitro pineapple (Ananas comosus L. Merr. var MD-2) as CAM or C3 is regulated by the environmental conditions’’. Plant Cell Reports, vol. 31, no. 4, 2 de diciembre de 2011, pp. 757-769, ISSN 0721-7714, 1432-203X, DOI 10.1007/s00299-011-1195-7.
10. Rodrigues, M. J. P.; Verdoodt, V.; Pasqual, M. y de Proft, M. ‘‘Impacts of photoautotrophic and photomixotrophic conditions on in vitro propagated Billbergia zebrina (Bromeliaceae)’’. Plant Cell, Tissue and Organ Culture (PCTOC), vol. 123, no. 1, 11 de julio de 2015, pp. 121-132, ISSN 0167-6857, 1573-5044, DOI 10.1007/s11240-015-0820-5.
11. Aragón, C. E.; Sánchez, C.; Gonzalez-Olmedo, J.; Escalona, M.; Carvalho, L. y Amâncio, S. ‘‘Comparison of plantain plantlets propagated in temporary immersion bioreactors and gelled medium during in vitro growth and acclimatization’’. Biologia Plantarum, vol. 58, no. 1, 16 de noviembre de 2013, pp. 29-38, ISSN 0006-3134, 1573-8264, DOI 10.1007/s10535-013-0381-6.
12. Malabadi, R. B.; Kumar, S. V.; Mulgund, G. S. y Nataraja, K. ‘‘Induction of somatic embryogenesis in Papaya (Carica papaya)’’. Research in Biotechnology, vol. 2, no. 5, 24 de octubre de 2011, pp. 40-55, ISSN 2229-791X.
13. Kumar, R. P.; Kumar, R. S. y Lokman, H. M. ‘‘Propagation of papaya (Carica papaya L.) cv. Shahi through in vitro culture’’. Bangladesh Journal of Botany, vol. 41, no. 2, 22 de enero de 2013, pp. 191-195, ISSN 2079-9926, 0253-5416, DOI 10.3329/bjb.v41i2.13448.
14. Cabrera, J. C.; Gómez, R. K.; Diosdado, E.; Hormaza, J. V.; Iglesias, R.; Gutiérrez, A. E. y González, S. J. Procedimiento de obtención de una mezcla de oligosacáridos pécticos estimuladora del enraizamiento vegetal. no. 22859, Inst. Oficina Cubana de la Propiedad Industrial, Cuba, 2003.
15. Hernández, M. M.; Suárez, L. y Valcárcel, M. ‘‘Empleo del Pectimorf en la micropropagación de Spathiphyllum sp’’. Cultivos Tropicales, vol. 30, no. 3, septiembre de 2009, pp. 56-58, ISSN 0258-5936.
16. Falcón, A. B. ‘‘Actividad enraizadora de una mezcla de oligogalacturónidos en pecíolos de violeta africana (Saintpaulia ionantha)’’. Cultivos Tropicales, vol. 28, no. 2, 2012, pp. 87–90, ISSN 0258-5936.
17. Posada, P. L.; Kosky, R. G. y Reyes, M. ‘‘Embriogénesis somática en Carica papaya L. var. Maradol rojo’’. Biotecnología vegetal, vol. 7, no. 3, 2007, pp. 131-138, ISSN 1609-1841.
18. Murashige, T. y Skoog, F. ‘‘A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures’’. Physiologia Plantarum, vol. 15, no. 3, 1 de julio de 1962, pp. 473-497, ISSN 1399-3054, DOI 10.1111/j.1399-3054.1962.tb08052.x.
19. Cardona, A. C.; Araméndiz, T. H. y Barrera, C. C. ‘‘Estimación del área foliar de papaya (Carica papaya L.) basada en muestreo no destructivo’’. Revista U.D.C.A Actualidad, Divulgación Científica, vol. 12, no. 1, junio de 2009, pp. 131-139, ISSN 0123-4226.
20. Arrieta, R. B. G.; Villegas, M. Á.; Hernández, B. A.; Rodríguez, M. M. de las N.; Ruiz, P. L. del M. y García, V. E. ‘‘Estomas y vigor de naranjo «valencia» injertado en portainjertos tolerantes al virus de la tristeza de los cítricos’’. Revista Fitotecnia Mexicana, vol. 33, no. 3, septiembre de 2010, pp. 257-263, ISSN 0187-7380.
21. Sánchez, D. M. y Aguirreolea, J. ‘‘Transpiración y control estomático’’. En: Fundamentos de Fisiología Vegetal, edit. McGraw Hill-Interamericana, Madrid, España, 2008, p. 620, ISBN 978-84-481-5168-3.
22. Hofman, P.; Haisel, D.; Komenda, J.; Vágner, M.; Tichá, I.; Schäfer, C. y Čapková, V. ‘‘Impact of in vitro Cultivation Conditions on Stress Responses and on Changes in Thylakoid Membrane Proteins and Pigments of Tobacco during ex vitro Acclimation’’. Biologia Plantarum, vol. 45, no. 2, junio de 2002, pp. 189-195, ISSN 0006-3134, 1573-8264, DOI 10.1023/A:1015180219628.
23. IBM Corporation. IBM SPSS Statistics [en línea]. versión 21, [Windows], Multiplataforma, edit. IBM Corporation, U.S, 2012, Disponible en: <http://www.ibm.com>.
24. Kong, S. S.; So, Y. P. y Kee, Y. P. ‘‘Sugar metabolism, photosynthesis, and growth of in vitro plantlets of Doritaenopsis under controlled microenvironmental conditions’’. In Vitro Cellular & Developmental Biology - Plant, vol. 49, no. 4, 10 de mayo de 2013, pp. 445-454, ISSN 1054-5476, 1475-2689, DOI 10.1007/s11627-013-9524-x.
25. Anandan, R.; Thirugnanakumar, S.; Sudhakar, D. y Balasubramanian, P. ‘‘In vitro organogenesis and plantlet regeneration of (Carica papaya L.)’’. Journal of Agricultural Technology, vol. 7, no. 5, 2011, pp. 1339–1348, ISSN 1686-9141.
26. Sekeli, R.; Abdullah, J. O.; Namasivayam, P.; Muda, P. y Abu Bakar, U. K. ‘‘Better rooting procedure to enhance survival rate of field grown Malaysian Eksotika papaya transformed with 1-Aminocyclopropane-1-carboxylic acid oxidase gene’’. ISRN Biotechnology, vol. 2013, 2012, ISSN 2090-9407, DOI 10.5402/2013/958945, [Consultado: 22 de marzo de 2016], Disponible en: <http://downloads.hindawi.com/journals/isrn.biotechnology/2013/958945.pdf>.
27. Cid, M.; González, O. J. L.; Lezcano, Y. y Nieves, N. ‘‘Influencia del Pectimorf sobre la calidad de la semilla artificial de caña de azúcar (Saccharum sp.)’’. Cultivos Tropicales, vol. 27, no. 1, 2006, pp. 31-34, ISSN 0258-5936.
28. González, P. L.; Vázquez, G. A.; Perrotta, L.; Acosta, A.; Scriven, S. A.; Herbert, R.; Cabrera, J. C.; Francis, D. y Rogers, H. J. ‘‘Oligosaccharins and Pectimorf® stimulate root elongation and shorten the cell cycle in higher plants’’. Plant Growth Regulation, vol. 68, no. 2, 3 de mayo de 2012, pp. 211-221, ISSN 0167-6903, 1573-5087, DOI 10.1007/s10725-012-9709-z.
29. Sáez, P. L.; Bravo, L. A.; Sáez, K. L.; Sánchez, O. M.; Latsague, M. I. y Ríos, D. G. ‘‘Photosynthetic and leaf anatomical characteristics of Castanea sativa: a comparison between in vitro and nursery plants’’. Biologia Plantarum, vol. 56, no. 1, 21 de enero de 2012, pp. 15-24, ISSN 0006-3134, 1573-8264, DOI 10.1007/s10535-012-0010-9.
30. Dominí, M. E. ‘‘Uso de biopreparados como promotores de enraizamientos en margullos de Ficus (Ficus benjamina)’’. Cultivos Tropicales, vol. 25, no. 3, 2004, pp. 45–48, ISSN 0258-5936.
Recibido: 18/12/2014
Aceptado: 30/10/2015
M.Cs. Laisyn Posada Pérez, Instituto de Biotecnología de las Plantas, Universidad Central “Marta Abreu” de Las Villas, arretera a Camajuaní, km 5,5, Santa Clara, Villa Clara, Cuba. Email: laisyn@ibp.co.cu