http://dx.doi.org/10.13140/RG.2.1.2060.0568
Efecto combinado de bajas dosis de rayos X y Biobras-16 en plantas de tomate (Solanum lycopersicum L.) cv. Vyta en condiciones de salinidad
Effect of the combined treatments of low X-rays doses and Biobras-16 on tomato (Solanum lycopersicum L.) cv. Vyta for salinity conditions
Yanelis Camejo-Serrano,I Ramiro Ramírez-Fernández,I Franklin Rivera-Pineda,II Maria C. González-Cepero,III Luis Licea-Castro,I Elia Porra-LeónI
IInstituto de Investigaciones Agropecuarias “Jorge Dimitrov”, carretera de Manzanillo, km. 16½, apartado Postal 2140, Bayamo, Granma, Cuba.
IIUniversidad Intercultural del Estado de México (UIEM), México.
IIIInstituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
RESUMEN
La salinidad es una de las condiciones adversas del medio que mayores estragos causa a los cultivos agrícolas. El objetivo del trabajo fue evaluar el efecto combinado de bajas dosis de rayos X y diferentes concentraciones de Biobras-16 en plantas de tomate, cultivadas en condiciones de salinidad. El experimento se desarrolló en el Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, en los años 2009 hasta el 2012. Se irradiaron semillas frescas de tomate cv. Vyta, con bajas dosis de rayos X (20, 25 y 30 Gy) y a los 15 días después del trasplante. Las plantas crecidas se asperjaron con dos concentraciones (1,5 y 2,0 mg L-1) de Biobras-16. Se emplearon dos sustratos, uno constituido por un suelo Vertisol salinizado con conductividad eléctrica de 6 dS m-1 del extracto de saturación y un suelo Vertisol no salino. Se observó que la mejor combinación de tratamientos fue 30 Gy+2,0 mg L-1, la cual incrementó significativamente (p≤0,05) la tolerancia a la salinidad, el rendimiento y sus componentes en las plantas de tomate.
Palabras clave: brasinosteroides, estrés salino, radioestimulación, tolerancia.
ABSTRACT
Salinity is one of the adverse environmental condition that cause high affectations to agricultural crops. The objective was to evaluate the combined effect of low doses of X-rays and different concentrations of Biobras-16 on tomato plants grown in saline conditions. The experiment was carried out at “Jorge Dimitrov” Agricultural Research Institute during the years 2009 to 2012. Fresh tomato seeds of variety cv. Vyta were exposed to X-ray doses of 20, 25 and 30 Gy and 15 days after transplantation the plants were sprayed with two concentrations (1,5 and 2,0 mg L-1) of Biobras-16. Two substrates were used, one consisting of a Vertisol soil salinized with electrical conductivity of 6 dS m-1 extract of soil saturation and Vertisol soil no salinized. The best combination of treatments was observed for 30 Gy +2,0 mg L-1, which increased significant (p≤0,05) the salt tolerance in plants, the tomato yield and its components.
Key words: brassinosteroids, saline stress, radiostimulation, tolerance.
INTRODUCCIÓN
Uno de los factores abióticos que afecta a los cultivos agrícolas, es la salinidad de los suelos. Aproximadamente 800 millones de hectáreas de la superficie terrestre se encuentra afectada por niveles de salinidad que, en muchos casos, superan la tolerancia de las especies tradicionales (1).
Este factor estresante provoca cuantiosas pérdidas en los rendimientos agrícolas de numerosos cultivos, incluyendo al tomate (2). Las principales afectaciones asociadas a la salinidad en las plantas son el déficit hídrico, el daño celular y el desbalance nutricional (3). La salinidad disminuye la fotosíntesis y aumenta el estrés oxidativo.
Todos estos trastornos implican una afectación en el crecimiento y desarrollo de los cultivos y por tanto una disminución del rendimiento (4, 5).
En los últimos años varios autores, han confirmado el efecto estimulante de las bajas dosis de rayos X en numerosas especies de plantas, corroborándose además, que su efectividad no depende solamente de la dosis integral de irradiación absorbida por el organismo, sino también de una serie de factores biológicos o ambientales interrelacionados que pueden potenciar o disminuir este efecto estimulante (6).
Además, se ha confirmado que la combinación de bajas dosis de radiaciones ionizantes con hormonas del crecimiento maximiza el efecto estimulante. En tal sentido, se estudió la respuesta de las plantas ante la combinación de las radiaciones ionizantes con algunas hormonas vegetales; observándose una respuesta sinérgica de las auxinas, giberelinas, citoquininas, ácido abscísico y etileno con las bajas dosis de rayos X (7).
Por otro parte, los brasinoesteroides han despertado un gran interés en la comunidad científica internacional, se ha demostrado que algunas formulaciones que tiene como ingrediente activo análogo de brasinoesteroides, como el Biobras-16 (BB-16), puede ser utilizado en la estimulación de los rendimientos agrícolas de cultivos de importancia económica (8–10).
Por tanto, el estudio del efecto combinado de las radiaciones ionizantes y los brasinoesteroides, considerados la sexta hormona del crecimiento vegetal reviste una gran importancia desde el punto de vista teórico y práctico; máxime si se considera que el posible efecto sinérgico de dicha hormona vegetal y las bajas dosis de rayos X pudieran inducir cierto grado de tolerancia y mitigar el efecto negativo de la salinidad en plantas de tomate.
El objetivo del este trabajo fue evaluar el efecto combinado de bajas dosis de rayos X aplicadas a las semillas y la aspersión de diferentes concentraciones de Biobras-16 en plantas de tomate.
MATERIALES Y MÉTODOS
La investigación, se desarrolló en áreas experimentales del Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, de la provincia Granma, Cuba, durante los años 2009-2012; enmarcados en los meses de noviembre a febrero, considerados óptimos para la producción de hortalizas.
En el experimento se emplearon las mejores combinaciones de diferentes concentraciones de Biobras-16 (BB-16) y bajas dosis de rayos X, evaluadas y seleccionadas en experimentos realizados con anterioridad (Tabla I). Las semillas fueron tratadas en horas de la mañana, en un equipo de rayos X, marca Philips, con un filtro de aluminio de 0,75 mm y un régimen de trabajo de 55 KV y 30 mA, una potencia de dosis de 11,47 Gy/min con temperatura de 24±1 °C.

En los experimentos se emplearon dos sustratos, uno constituido por un suelo Vertisol de tipo salinizado con predominio de sales neutras como cloruros y sulfatos de sodio, calcio y magnesio, pH< 7,6 para una concentración de sales solubles totales de 3 565 ppm y valores medios de conductividad eléctrica en el extracto de saturación de 6 dS m-1, por lo que presenta un grado de salinidad S3 según la Norma Cubana, 2009, donde se sembraron los tratamientos (T0, T1, T2, T3, ); y otro compuesto por un suelo Vertisol no salino (11), donde se plantaron los tratamientos y el control.
Las plántulas procedentes de los semilleros se trasplantaron a condiciones semicontroladas en el mes de diciembre, cuando reunieron los requisitos necesarios establecidos, según el Manual técnico para organopónicos, huertos intensivos y organoponía semiprotegida (12). Se utilizaron macetas de 23 cm de ancho por 29 cm de profundidad; se trasplantaron 20 plántulas por réplica, siguiendo un diseño completamente aleatorizado, con cuatro replicas por tratamiento.
Durante el periodo de floración (15 días después del trasplante), fueron realizadas las aplicaciones de las distintas concentraciones del BB-16 (Tabla I) a razón de 2 mL por planta. Las hojas fueron asperjadas durante las primeras horas de la mañana hasta que estuvieron completamente mojadas. Se utilizó una mochila de 16 litros con un aspersor de boquilla cónica, previamente calibrada.
Se evaluaron los siguientes indicadores: longitud y diámetro del tallo y raíz, área foliar, porcentaje de materia seca, número de racimos por planta, número de frutos por racimo, diámetro polar y ecuatorial (cm), masa promedio de los frutos (g) y rendimiento por planta (kg planta-1). Las evaluaciones del crecimiento se realizaron a los 20 días después del trasplante, y las evaluaciones del rendimiento y sus componentes al completar el ciclo del cultivo, en 80 plantas por tratamiento. Los datos presentados constituyen la media de los tres años, ya que no existieron diferencias significativas entre ellos.
Con los datos obtenidos se calculó la tolerancia de las plantas a la salinidad, mediante la fórmula descrita (13):
TS=100(IS/IC)
donde:
TS: tolerancia a la salinidad
IS: media de los indicadores evaluados en condiciones de salinidad
IC: media de los indicadores evaluados en condiciones normales
Los datos del crecimiento, del rendimiento y sus componentes, así como de la tolerancia a la salinidad, se procesaron mediante un análisis de varianza de clasificación simple, y al comprobarse diferencias significativas se utilizó la prueba de comparación múltiple de medias de Tukey, para una probabilidad de error del 5 %.
RESULTADOS Y DISCUSIÓN
Al evaluar el efecto combinado de bajas dosis de rayos X y BB-16 en plantas de tomate, en condiciones de salinidad, se observó una estimulación significativa (p≤0,05) en algunos indicadores de crecimiento (Tabla II), que permitió minimizar el efecto del estrés, al mostrar una respuesta similar a las plántulas cultivadas en condiciones normales.

La mejor respuesta, de manera general, se obtuvo en el tratamiento T2, el cual estimuló significativamente todos los indicadores del crecimiento evaluados.
La longitud del tallo (LT), incrementó significativamente (p≤0,05), excepto el tratamiento T3, el cual no mostró diferencias con relación al control. La mayor respuesta en este indicador se observó en los tratamientos T1 y T2, el cual se corresponde con los tratamientos combinado de 20 Gy y 2,0 mg L-1 y 30 Gy y 2,0 mg L-1.
En relación con el indicador diámetro del tallo (DT) se constataron diferencias significativas (p≤0,05) con respecto al control, en el tratamiento T2; mientras que la longitud y diámetro de la raíz (LR y DR) se estimularon significativamente todos los tratamientos pero no hubo diferencias significativas entre ellos.
El área foliar (AF) mostró estimulación significativa (p≤0,05) con los tres tratamientos, observándose los mayores incrementos coincidentemente en el tratamiento T2.
Un incremento de este indicador pudiera favorecer la asimilación de energía solar en el proceso de fotosíntesis e implicar una mayor producción de fotoasimilados, lo que contribuye mayormente al crecimiento y desarrollo de las plantas (14).
El incremento de estos indicadores del crecimiento ha sido informado por otros autores, al emplear estos métodos. Se ha evidenciado una respuesta del crecimiento en plantas de lechuga (Lactuca sativa L.) con la aspersión foliar de Biobras-16 incrementándose tanto el desarrollo foliar como el crecimiento radical (15).
Por otro lado, en plántulas de tomate de la variedad Amalia, se observó un incremento de la longitud de la raíz, al aplicar Biobras-16 en un medio salino; lo que sugiere la capacidad de este análogo de brasinoesteroide de inducir la activación del mecanismo de tolerancia al NaCl en las raíces (16).
Se ha encontrado además, una estimulación altamente significativa (30 %) del indicador área foliar, en cuatro variedades de tomates, al ser tratadas con bajas dosis de rayos XA.
Otros autores observaron un incremento de los indicadores del crecimiento en plantas de Arabidopsis thaliana Landsberg (Ler) al aplicar bajas dosis de rayos gamma (100 Gy), estimulándose además, el número de flores y silicuas (17).
Al respecto, se observó un incremento de la longitud de la raíz en plantas de Pterocarpus marsupium Roxbcon respecto al control, al aplicar bajas dosis de radiación gamma (18).
En el porcentaje de materia seca se observó diferencias significativas (p≤0,05) de los tratamientos T1 y T2 con respecto al control, mientras que el tratamiento T3 no mostró diferencias significativas (Tabla III). Este es un aspecto de gran importancia ya que se considera por otros autores un indicador preciso de tolerancia en otras especies, tomate (19) y en trigo (20).
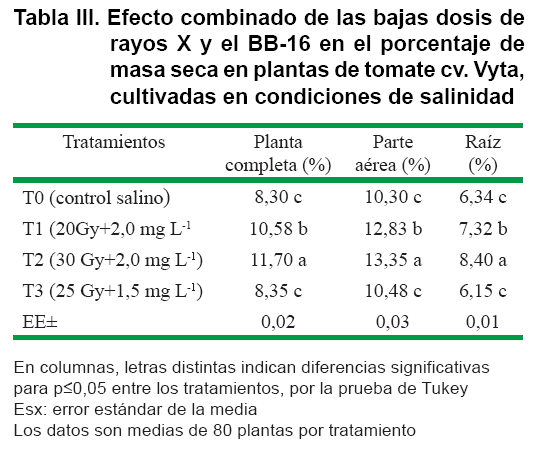
Similares resultados se informaron, al aplicar la aspersión foliar de 24-epibrasinolida en el cultivo del tomate, observándose un incremento de la acumulación de la biomasa y de la actividad fotosintética (21, 22).
Otros autores observaron incrementos en los indicadores del crecimiento, materia seca y longitud de la raíz, en plantas de Arabidopsis thaliana al estimular con dosis de 10 Gy de rayos X (23).
Estos resultados demuestran la efectividad al combinar las bajas dosis de rayos X y la aspersión del BB-16 en el incremento del crecimiento en condiciones de salinidad en plantas de tomate cv. Vyta.
Esta respuesta pudiera estar relacionada, por una parte, con la hormesis inducida por las bajas dosis de radiaciones ionizantes que se manifiesta con una activación e incremento de los procesos oxidativos; como consecuencia de las cuales aumenta el metabolismo químico general de las células, todo lo cual conlleva a transformaciones energéticas en las plantas que se reflejan durante su ciclo vital (24).
De acuerdo con esta teoría se ha señalado que al producirse un aumento en los procesos oxidativos se estimulan determinadas funciones vitales del organismo vegetal, relacionadas con variaciones en la actividad de ciertas enzimas, en el estado de agregación del protoplasma y en sus propiedades coloidales, que se expresan en un crecimiento generalizado del metabolismo químico (25).
Por otra parte, los brasinoesteroides también poseen la capacidad de acelerar el crecimiento y la maduración de las plantas; además los efectos inducidos por los brasinoesteroides no pueden ser considerados en forma aislada, ya que estos compuestos interactúan con otros reguladores endógenos del crecimiento vegetal y son muy sensibles a señales ambientales particularmente con la calidad de la irradiación solar, respuesta típica de otras hormonas vegetales (26).
En este sentido la actividad de la hormona pudiera estar influenciada con las bajas dosis de rayos X y generar un efecto estimulante en los indicadores del crecimiento en las plantas de tomate en condiciones de salinidad, lo que conlleva a incrementos significativos en el rendimiento.
Se observó además, una estimulación significativa (p≤0,05) del rendimiento y sus componentes (Figura). La respuesta de las variables número de frutos y racimos por planta mostró diferencias significativas (p≤0,05) entre los tratamientos aplicados y el control; se destacan T1 y T2 en el primer indicador y T2 en el segundo.
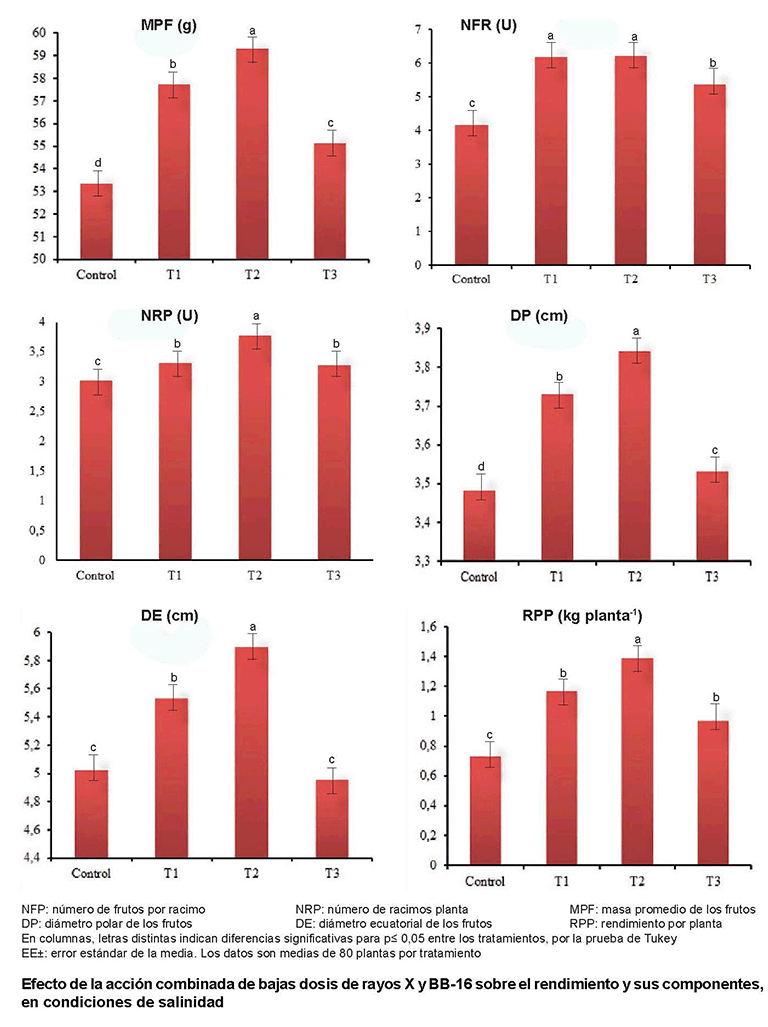
El incremento observado en el número de frutos por planta se considera un aspecto de gran importancia ya que esta variable es un indicador directo del rendimiento agrícola y de adaptación a las condiciones edafoclimáticas del cultivo (27); por lo que, otros autores recomiendan utilizarla para seleccionar variedades adaptadas a las condiciones adversas del medio (28).
No obstante al efecto negativo de la salinidad en las plantas, la masa promedio de los frutos también se estimuló significativamente (p≤0,05) en todos los tratamientos combinados (T1-T3), y coincide el T2 como el de mejor respuesta, al lograr valores significativos de estimulación de 60 g en este indicador. Este resultado pudiera indicar la posibilidad de utilizar el efecto combinado de las bajas dosis de rayos X y el Biobras-16 en la inducción de tolerancia a la salinidad en este cultivo.
En las variables diámetro ecuatorial y polar de los frutos se constataron incrementos significativos (p≤0,05) de estimulación. El tratamiento T2 mostró los mayores valores respecto al control. Excepto T3 en el indicador diámetro ecuatorial, el resto de los tratamientos combinados indujeron una respuesta positiva ante las condiciones de salinidad.
En el rendimiento por planta se observó la presencia de valores significativos (p≤0,05) de estimulación en todos los tratamientos aplicados (T1-T3), con valores que oscilaron entre 0,97 y 1,39 kg planta-1.
Estos resultados corroboran lo observado por otros autores que indican la factibilidad de utilizar, tanto las radiaciones ionizantes (29) como los brasinoesteroides (30) para incrementar la tolerancia de las plantas al estrés abiótico. En este sentido, se logró en la variedad de tomate Campbell–28, una estimulación del rendimiento del 40 %, al tratar sus semillas con bajas dosis de rayos X, cultivadas en medio salino (31). Se encontró, además, un incremento significativo en los indicadores número de racimos por planta, número de frutos por planta y el rendimiento, con dosis de 20 y 25 Gy en la variedad INCA-9-1 y en los mutantes Maybel y DomiB.
De acuerdo con los autores citados, esta respuesta puede ser atribuida a que ambos métodos activan de forma general una serie de reacciones de defensa adaptativas del organismo vegetal contra los daños producidos por las condiciones adversas del medio. Se plantea que estos estimuladores provocan una disminución en la penetrabilidad del protoplasma y frenan la acumulación de iones tóxicos en las plantas, lo que conlleva a un incremento de la tolerancia al estrés (30, 31).
Un análisis teórico del tema sugiere que tanto los rayos X como los brasinoesteroides incrementan significativamente la actividad de las enzimas ARN y ADN polimerasas, la síntesis del ARN, ADN y proteínas. Bajo la influencia de ambos estimulantes, los cambios en las actividades enzimáticas aparentemente afectan el metabolismo de los ácidos nucleicos, de tal forma que los niveles de ARN, ADN y proteínas acumulados en el tejido se incrementan durante el crecimiento (32, 33).
Al analizar la respuesta a la salinidad en plantas de tomate de la variedad Vyta, expresado mediante el índice relativo de tolerancia, se constató un efecto diferenciado en los indicadores del rendimiento y sus componentes. Los tres tratamientos combinados con rayos X y Biobras-16 (Tabla IV) igualan o superan significativamente el índice de tolerancia relativa mostrado por el control.
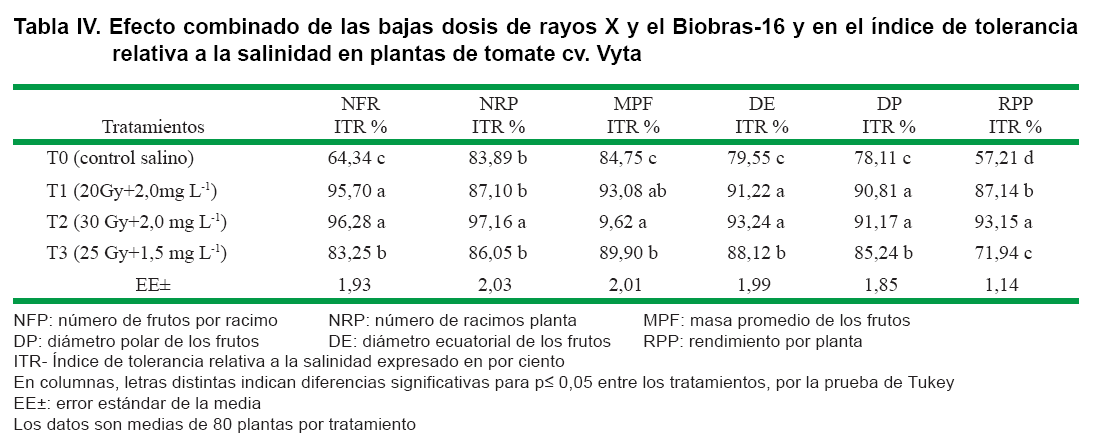
El cálculo del índice de tolerancia a la salinidad en el indicador NFR señala el efecto combinado de ambos métodos estimulantes al incrementar significativamente la tolerancia de Vyta; que alcanzó valores máximos en los tratamientos T1 y T2, respectivamente.
El NRP se expresó de forma similar al anterior indicador, con respecto al grado de tolerancia. Coincide el tratamiento T2 con el de mejor respuesta en la inducción de tolerancia, el cual mostró valores de 97,16 %. Por otro lado, las plantas provenientes del tratamiento T1 manifestaron una respuesta similar a T3.
La MPF se afectó en menor medida por la salinidad; no obstante, los índices de tolerancia aumentaron significativamente con los tratamientos aplicados, de tal manera que T2 mostró las mejores respuestas en estas condiciones de estrés.
En los índices de tolerancia observados en los DP y DE de los frutos se constató una respuesta similar. Se destacan, por el incremento en dichos índices, los tratamientos T1 y T2, aunque T3 también mostro diferencias significativas con el control.
En el indicador RPP los tres tratamientos combinados estimulantes, al parecer, indujeron un alto grado de tolerancia, pues se destaca coincidentemente, el tratamiento T2, que alcanzó un índice de 93,15 %.
Es preciso señalar que la salinidad afecta el rendimiento y sus componentes; al respecto otros autores, coinciden en que tales afectaciones en las plantas cultivadas, bajo condiciones de estrés, son debidas a las variaciones que se producen en la concentración y en la relación de las hormonas estimuladoras e inhibidoras del crecimiento y desarrollo, destacando que este desbalance hormonal juega un rol importante en la regulación de la respuesta de las plantas a la salinidad, controlando fundamentalmente los procesos de síntesis de compuestos orgánicos (34).
Los valores obtenidos en los indicadores de tolerancia a la salinidad, en el rendimiento y sus componentes, muestran la capacidad del análogo de barsinoesteroide y los rayos X en la inducción de tolerancia; más si se tiene en cuenta que desde el punto de vista agronómico, la tolerancia a la salinidad se define como la habilidad de las plantas de sobrevivir y producir rendimientos económicos en condiciones de estrés, y se expresa como la relación entre el rendimiento de una variedad en condiciones salinas, con respecto a su rendimiento, en condiciones normales.
Los brasinoesteroides son considerados potentes reguladores del crecimiento vegetal de naturaleza esteroidal. Estas hormonas tienen efectos pleiotrópicos como son: estimulación del alargamiento celular y de la desdiferenciación de protoplastos, regeneración de la pared celular, regulación de la diferenciación de elementos traqueales e incremento de la biomasa y del rendimiento en diferentes especies de plantas (35).
Diversos autores, hacen referencia también al efecto protector de los brasinoesteroides ante diferentes condiciones de estrés abiótico, como altas y bajas temperaturas, sequía y salinidad; además, disminuyen los efectos del estrés causado por la falta de nutrientes y por exceso de metales pesados y también incrementan la resistencia a herbicidas y a agentes patógenos (36).
Por otro lado, las radiaciones ionizantes, de manera general, a bajas dosis son consideradas potentes agentes estimulantes, y con mayor eficiencia los rayos X, teniendo en cuenta su menor longitud de onda que disminuye la probabilidad de provocar o inducir mutaciones (29, 37).
Numerosos autores informaron las ventajas más significativas de la radioestimulación o radiohormesis, relacionadas con incrementos en los rendimientos agrícolas (10-40 %), la germinación de las semillas, el contenido de carotenos y vitamina C en algunas hortalizas y de proteínas y grasas en cereales (38), también se han mostrado su eficacia en el incremento de la tolerancia a factores abióticos del medio como la salinidad (31).
También, otros autores observaron un efecto estimulante y protector ante factores bióticos y abióticos del medio y un incremento en la actividad de las enzimas antioxidantes, así como un incremento en la activación de la Poli (ADN-ribosa) polimerasa, enzima reguladora de la respuesta al estrés, como señal de tolerancia (39, 40).
A la vez, los brasinoesteroides poseen una incidencia marcada en el crecimiento de las plantas en condiciones de salinidad. Experimentos realizados demostraron la efectividad del análogo espirostánico de brasinoesteroide (BB-16) en revertir la inhibición que provocó la salinidad en el crecimiento de las plántulas de dos genotipos de arroz (41).
CONCLUSIONES
Los tratamientos combinados con bajas dosis de rayos X y Biobras-16 pueden ser utilizados para lograr un efecto estimulante en la tolerancia a la salinidad en plantas de tomate cv. Vyta e incrementos significativos en el crecimiento, rendimiento y sus componentes.
Notas al pie
ARamírez, R. Efecto del tratamiento de semillas con dosis estimulantes de rayos X en el cultivo del tomate (Lycopersicum esculentum Mill.). [Tesis de Doctorado], Instituto Nacional de Ciencias Agrícolas, La Habana, Cuba, 2006, 130 p.
BOjeda, C. M. Incremento de la tolerancia a la salinidad en plantas de tomate (Solanum lycopersicum L), mediante el tratamiento de semillas con radiaciones ionizantes. [Tesis de Maestría], Facultad de Ciencias Agrícola, Universidad de Granma, Bayamo, Cuba, 2009, 69 p.
BIBLIOGRAFÍA
1.FAO. Salt-affected soils [en línea]. Portal de Suelos de la FAO, 2016, [Consultado: 4 de mayo de 2016], Disponible en: <http://www.fao.org/soils-portal/manejo-del-suelo/manejo-de-suelos-problematicos/suelos-afectados-por-salinidad/more-information-on-salt-affected-soils/es/>.
2. Li, C.; Chang, P. P.; Ghebremariam, K. M.; Qin, L. y Liang, Y. ‘‘Overexpression of tomato SpMPK3 gene in Arabidopsis enhances the osmotic tolerance’’. Biochemical and Biophysical Research Communications, vol. 443, no. 2, 10 de enero de 2014, pp. 357-362, ISSN 0006-291X, DOI 10.1016/j.bbrc.2013.11.061.
3. Talaat, N. B. y Shawky, B. T. ‘‘Protective effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) plants exposed to salinity’’. Environmental and Experimental Botany, vol. 98, febrero de 2014, pp. 20-31, ISSN 0098-8472, DOI 10.1016/j.envexpbot.2013.10.005.
4. Dudhane, M. P.; Borde, M. Y. y Jite, P. K. ‘‘Effect of arbuscular mycorrhizal fungi on growth and antioxidant activity in Gmelina arborea Roxb. under salt stress condition’’. Notulae Scientia Biologicae, vol. 3, no. 4, 2011, p. 71, ISSN 2067-3265.
5. Talaat, N. B. y Shawky, B. T. ‘‘24-Epibrassinolide ameliorates the saline stress and improves the productivity of wheat (Triticum aestivum L.)’’. Environmental and Experimental Botany, vol. 82, octubre de 2012, pp. 80-88, ISSN 0098-8472, DOI 10.1016/j.envexpbot.2012.03.009.
6. de Micco, V.; Arena, C.; Pignalosa, D. y Durante, M. ‘‘Effects of sparsely and densely ionizing radiation on plants’’. Radiation and Environmental Biophysics, vol. 50, no. 1, 27 de noviembre de 2010, pp. 1-19, ISSN 0301-634X, 1432-2099, DOI 10.1007/s00411-010-0343-8.
7. Kumar, V. y Kumar, D. ‘‘Synergistic effect of the combined treatment with ray X and phytohormones in tomato’’. Indian Journal of Agricultural Sciences, vol. 58, no. 4, 1987, pp. 313-314, ISSN 0019-5022.
8. Alarcón, Z. A.; Barreiro, E. P. y Díaz, S. Y. ‘‘Efecto del Biobras-16 y el Fitomas-E en algunos indicadores del crecimiento y el rendimiento del tomate (Solanum Lycopersicum, Lin) variedad «Vyta»’’. Revista Granma Ciencia, vol. 16, no. 1, 2012, pp. 3-10, ISSN 1027-975X.
9. Serna, M.; Hernández, F.; Coll, F. y Amorós, A. ‘‘Brassinosteroid analogues effect on yield and quality parameters of field-grown lettuce (Lactuca sativa L.)’’. Scientia Horticulturae, vol. 143, 16 de agosto de 2012, pp. 29-37, ISSN 0304-4238, DOI 10.1016/j.scienta.2012.05.019.
10. Serna, L. ‘‘The role of brassinosteroids and abscisic acid in stomatal development’’. Plant Science, vol. 225, agosto de 2014, pp. 95-101, ISSN 0168-9452, DOI 10.1016/j.plantsci.2014.05.017.
11. Hernández, A.; Ascanio, M. O.; Morales, M. y Cabrera, A. ‘‘Correlación de la Nueva Versión de Clasificación Genética de los Suelos de Cuba, con clasificaciones internacionales (Soil Taxonomy y FAO-UNESCO) y clasificaciones nacionales (2da Clasificación Genética y Clasificación de series de suelos)’’. En: VI Congreso Nacional de la Ciencia del Suelo, edit. Instituto de Suelos, Ministerio de la Agricultura y Sociedad Cubana de la Ciencia del Suelo, La Habana, Cuba, 2006, p. 62, ISBN 959-7023-35-0.
12. Rodríguez, A.; Companioni, N.; Peña, E.; Cañet, F.; Fresneda, J.; Estrada, J. y Rey, R. Manual técnico para organopónicos, huertos intensivos y organoponía semiprotegida. 6.a ed., edit. Instituto de Investigaciones Fundamentales en Agricultura Tropical (INIFAT), Cuba, 2007, ISBN 978-959-246-030-2.
13. González, L. M.; López, R. C.; Fonseca, I. y Ramírez, R. ‘‘Crecimiento, frecuencia estomática, rendimiento de MS y acumulación de iones en nueve especies de leguminosas pratenses cultivadas en condiciones salinas’’. Pastos y Forrajes, vol. 23, no. 4, 2000, pp. 299-308, ISSN 2078-8452.
14. Bhargava, Y. R. y Khalatkar, A. S. ‘‘Improve performance of Tectona grandis seeds with gamma irradiation’’. Acta Horticulturae, no. 215, octubre de 1987, pp. 51-54, ISSN 0567-7572, 2406-6168, DOI 10.17660/ActaHortic.1987.215.7.
15. Terry, A. E.; Ruiz, P. J.; Tejeda, P. T.; Reynaldo, E. I. y Díaz, de A. M. M. ‘‘Respuesta del cultivo de la lechuga (Lactuca sativa L.) a la aplicación de diferentes productos bioactivos’’. Cultivos Tropicales, vol. 32, no. 1, marzo de 2011, pp. 28-37, ISSN 0258-5936.
16. Reyes, G. Y.; Rosabal, A. L.; Martínez, G. L.; Mazorra, M. L. M. y Núñez, V. M. ‘‘Efecto de los brasinoesteroides y un inhibidor de su biosíntesis en plántulas de dos variedades de tomate sometidas a estrés salino’’. Cultivos Tropicales, vol. 35, no. 1, marzo de 2014, pp. 25-34, ISSN 0258-5936.
17. Kim, D. S.; Kim, J. B.; Goh, E. J.; Kim, W. J.; Kim, S. H.; Seo, Y. W.; Jang, C. S. y Kang, S. Y. ‘‘Antioxidant response of Arabidopsis plants to gamma irradiation: Genome-wide expression profiling of the ROS scavenging and signal transduction pathways’’. Journal of Plant Physiology, vol. 168, no. 16, 1 de noviembre de 2011, pp. 1960-1971, ISSN 0176-1617, DOI 10.1016/j.jplph.2011.05.008.
18. Chandrashekar, K. R. A. ‘‘Gamma sensitivity of forest plants of Western Ghats’’. Journal of Environmental Radioactivity, vol. 132, junio de 2014, pp. 100-107, ISSN 0265-931X, DOI 10.1016/j.jenvrad.2014.02.006.
19. Almasoum, A. A. ‘‘Effect of planting depth on growth and productivity of tomato es using drip irrigation with semi saline water’’. Acta Horticulturae, no. 537, octubre de 2000, pp. 773-778, ISSN 0567-7572, 2406-6168, DOI 10.17660/ActaHortic.2000.537.92.
20. Argentel, M. L.; Garatuza, P. J.; Yépez, G. E. A. y de los Santos-Villalobos, S. ‘‘Evaluación de la tolerancia de variedades mexicanas de trigo a la salinidad, a través de indicadores fisiológicos, bioquímicos y agronómicos, cultivadas en Cuba en condiciones de campo’’. Cultivos Tropicales, vol. 37, no. 1, marzo de 2016, pp. 91-101, ISSN 0258-5936.
21. Ahammed, G. J.; Gao, C. J.; Ogweno, J. O.; Zhou, Y. H.; Xia, X. J.; Mao, W. H.; Shi, K. y Yu, J. Q. ‘‘Brassinosteroids induce plant tolerance against phenanthrene by enhancing degradation and detoxification in Solanum lycopersicum L.’’. Ecotoxicology and Environmental Safety, vol. 80, 1 de junio de 2012, pp. 28-36, ISSN 0147-6513, DOI 10.1016/j.ecoenv.2012.02.004.
22. Ahammed, G. J.; Yuan, H. L.; Ogweno, J. O.; Zhou, Y. H.; Xia, X. J.; Mao, W. H.; Shi, K. y Yu, J. Q. ‘‘Brassinosteroid alleviates phenanthrene and pyrene phytotoxicity by increasing detoxification activity and photosynthesis in tomato’’. Chemosphere, vol. 86, no. 5, febrero de 2012, pp. 546-555, ISSN 0045-6535, DOI 10.1016/j.chemosphere.2011.10.038.
23. Gicquel, M.; Esnault, M. A.; Jorrín, N. J. V. y Cabello, H. F. ‘‘Application of proteomics to the assessment of the response to ionising radiation in Arabidopsis thaliana’’. Journal of Proteomics, vol. 74, no. 8, 12 de agosto de 2011, pp. 1364-1377, ISSN 1874-3919, DOI 10.1016/j.jprot.2011.03.025.
24. Kuzin, A. ‘‘The key mechanisms of radiation hormesis’’. Izvestiia Akademii nauk. Seriia biologicheskaia / Rossiiskaia akademiia nauk, no. 6, diciembre de 1992, pp. 824-832, ISSN 1026-3470.
25. Calabrese, E. J. y Baldwin, L. A. ‘‘Chemical hormesis: its historical foundations as a biological hypothesis’’. Human & Experimental Toxicology, vol. 19, no. 1, 1 de enero de 2000, pp. 2-31, ISSN 0960-3271, 1477-0903, DOI 10.1191/096032700678815585.
26. Barbafieri, M. y Tassi, E. ‘‘Brassinosteroids for phytoremediation application’’ [en línea]. En: eds. Hayat S. y Ahmad A., Brassinosteroids: A Class of Plant Hormone, edit. Springer Netherlands, 2011, pp. 403-437, ISBN 978-94-007-0188-5, [Consultado: 23 de marzo de 2016], Disponible en: <http://link.springer.com/chapter/10.1007/978-94-007-0189-2_16>.
27. Rodríguez, J.; Álvarez, M.; Moya, C.; Plana, D.; Dueñas, F.; Lescay, E. y Rodríguez, S. ‘‘Identificación de progenitores de tomate (Solanum lycopersicum) para la obtención de híbridos F1 adaptados a las condiciones de Cuba’’. Cultivos Tropicales, vol. 29, no. 3, septiembre de 2008, pp. 69-72, ISSN 0258-5936.
28. Plana, D.; Moya, C.; Álvarez, M.; Dueñas, F. y Pino, M. de los A. ‘‘Agricultores urbanos participando en la selección de variedades de tomate’’. En: XIV Congreso Científico del INCA, edit. INCA, La Habana, Cuba, 2004, p. 148.
29. Arena, C.; de Micco, V.; Aronne, G.; Pugliese, M.; Virzo, de S. A. y de Maio, A. ‘‘Response of Phaseolus vulgaris L. plants to low-let ionizing radiation: Growth and oxidative stress’’. Acta Astronautica, vol. 91, octubre de 2013, pp. 107-114, ISSN 0094-5765, DOI 10.1016/j.actaastro.2013.05.013.
30. Vriet, C.; Russinova, E. y Reuzeau, C. ‘‘Boosting Crop Yields with Plant Steroids’’. The Plant Cell, vol. 24, no. 3, 1 de marzo de 2012, pp. 842-857, ISSN 1532-298X, DOI 10.1105/tpc.111.094912.
31. González, L. M.; Chávez, L.; Ramírez, R. y Camejo, Y. ‘‘Incremento de la tolerancia a la salinidad en plántulas de tomate (Lycopersicon esculentum Mill), mediante el tratamiento de semillas con rayos X’’. Alimentaria, no. 339, 2002, pp. 109-112, ISSN 0300-5755.
32. Esnault, M. A.; Legue, F. y Chenal, C. ‘‘Ionizing radiation: Advances in plant response’’. Environmental and Experimental Botany, vol. 68, no. 3, mayo de 2010, pp. 231-237, ISSN 0098-8472, DOI 10.1016/j.envexpbot.2010.01.007.
33. Choudhary, S. P.; Yu, J. Q.; Yamaguchi, S. K.; Shinozaki, K. y Tran, L. S. P. ‘‘Benefits of brassinosteroid crosstalk’’. Trends in Plant Science, vol. 17, no. 10, octubre de 2012, pp. 594-605, ISSN 1360-1385, DOI 10.1016/j.tplants.2012.05.012.
34. Syvertsen, J. P. y Garcia, S. F. ‘‘Multiple abiotic stresses occurring with salinity stress in citrus’’. Environmental and Experimental Botany, vol. 103, julio de 2014, pp. 128-137, ISSN 0098-8472, DOI 10.1016/j.envexpbot.2013.09.015.
35. Hasan, S. A.; Hayat, S. y Ahmad, A. ‘‘Brassinosteroids protect photosynthetic machinery against the cadmium induced oxidative stress in two tomato cultivars’’. Chemosphere, vol. 84, no. 10, septiembre de 2011, pp. 1446-1451, ISSN 0045-6535, DOI 10.1016/j.chemosphere.2011.04.047.
36. Núñez, V. M.; Reyes, G. Y.; Rosabal, A. L. y Martínez, G. L. ‘‘Análogos espirostánicos de brasinoesteroides y sus potencialidades de uso en la agricultura’’. Cultivos Tropicales, vol. 35, no. 2, junio de 2014, pp. 34-42, ISSN 0258-5936.
37. Iglesias, L.; Octavio, P. y Bello, J. ‘‘Current importance and potential use of low doses of gamma radiation in forest species’’. En: ed. Adrovic F., Gamma Radiation, edit. In Tech Publishing, Croatia, 2012, pp. 978-983, ISBN 978-953-51-0316-5.
38. Vasilevski, G. ‘‘Perspectives of the application of biophysical methods in sustainable agriculture’’. Bulgarian Journal of Plant Physiology, vol. 29, no. 3, 2003, pp. 179–186, ISSN 1312-8213.
39. Arena, C.; de Micco, V. y de Maio, A. ‘‘Growth alteration and leaf biochemical responses in Phaseolus vulgaris exposed to different doses of ionising radiation’’. Plant Biology, vol. 16, 1 de enero de 2014, pp. 194-202, ISSN 1438-8677, DOI 10.1111/plb.12076.
40. de Micco, V.; Arena, C. y Aronne, G. ‘‘Anatomical alterations of Phaseolus vulgaris L. mature leaves irradiated with X-rays’’. Plant Biology, vol. 16, 1 de enero de 2014, pp. 187-193, ISSN 1438-8677, DOI 10.1111/plb.12125.
41. Núñez, V. M.; Reyes, G. Y.; Rosabal, A. L.; Martínez, L.; González, C. M. C. y Pieters, A. ‘‘Brasinoesteroides y sus análogos estimulan el crecimiento de plántulas de dos genotipos de arroz (Oryza sativa L.) en medio salino’’. Cultivos Tropicales, vol. 34, no. 1, marzo de 2013, pp. 74-80, ISSN 0258-5936.
Recibido: 18/05/2015
Aceptado: 22/01/2016
Yanelis Camejo Serrano, Instituto de Investigaciones Agropecuarias “Jorge Dimitrov”, carretera de Manzanillo, km. 16½, apartado Postal 2140, Bayamo, Granma, Cuba. Email: ycamejo@dimitrov.cu