Rizobios residentes en la rizosfera de plantas de arroz (Oryza sativa L.) cultivar INCA LP-5
Residents rhizobia in the rhizosphere of rice plants (Oryza sativa L.) cultivar inca lp-5
M.Cs. Ionel Hernández Forte, Dra.C. María Caridad Nápoles García
Instituto Nacional de Ciencias Agrícolas (INCA), Gaveta postal No.1, San José de las Lajas. Mayabeque, Cuba. CP 32700.
Resumen
El objetivo del presente trabajo fue aislar rizobios de la rizosfera de plantas de arroz cultivar INCA LP-5. El aislamiento se realizó a partir del suelo rizosférico y del rizoplano de plantas de arroz cultivadas en suelo Gley Nodular Ferruginoso Petroférrico. Se determinó la posible distribución taxonómica de los aislados mediante el estudio de sus características morfo-culturales y fisiológicas y se realizaron bioensayos de nodulación in vitro en Macroptilium atropurpureum (DC.) Urb. (siratro). Se obtuvieron 14 aislados con características culturales y morfológicas similares a las descritas en los rizobios, de los cuales el 28,57 % se obtuvieron del suelo rizosférico y el 71,43 % del rizoplano de las plantas de arroz. Todos los aislados resultaron negativo a la prueba de la cetolactasa y produjeron ácido. Un aislado tuvo un crecimiento moderado sobre el medio de cultivo mientras que el resto crecieron más rápidamente. Once aislados provenientes de la rizosfera de las plantas de arroz produjeron nódulos efectivos en las plantas de siratro. Se identificó un aislado como posible miembro del género Mesorhizobium y diez en la familia Rhizobiaceae. Cuatro aislados presentaron una elevada efectividad del proceso de nodulación y uno de ellos incrementó significativamente la masa seca radical. Los resultados obtenidos en esta investigación constituyen la primera evidencia en Cuba donde se aborda la interacción de los rizobios con el cultivo del arroz.
Palabras clave: aislamiento, Rhizobiaceae, rizobacterias.
Abstract
The objective of this work was to isolate rhizobia from the rhizosphere of rice plants cultivar INCA LP-5. Rhizobia isolation from rhizospheric soil and rhizoplane of rice plants growing in Petroferric Ferruginous Nodular Gley soil was conducted. The possible taxonomic distribution of the isolates was determinated throught the study of their morpho-cultural and physiological characteristics. In vitro nodulation assays were also performed in Macroptilium atropurpureum (DC.) Urb. (siratro). Fourteen isolates showed similar cultural and morphological characteristics to rhizobia, of which 28,57 % were obtained from the rhizospheric soil and 71,43 % from rhizoplane of rice plants. All isolates were negative to cetolactase test and produced acid. An isolated showed moderate growth on the culture medium while the rest grew faster. Eleven rhizospherics isolated from rice plants produced effective nodules in siratro plants. A possible member of the Mesorhizobium genera and ten possible members of Rhizobiaceae family were identified. Four isolates showed a high effectiveness of the nodulation process in siratro plants and one of them significantly increased the root dry mass. The results obtained in this study provide the first evidence in Cuba where interaction of rhizobia-rice is developed.
Key words: isolation, Rhizobiaceae, rizobacteria.
INTRODUCCIÓN
El arroz es uno de los cultivos de mayor importancia para la alimentación humana, con una producción mundial de 478 millones de toneladasA. En América Latina, Cuba constituye uno de los países de mayor consumo per cápita, con alrededor de 60 kg anualesB. El cultivar de arroz INCA LP-5, obtenido en el Instituto Nacional de Ciencias Agrícolas, es uno de los de mayor distribución en el país por sus elevados rendimientos y por su resistencia a Pyricularia grisea, principal patógeno del cultivo (1).
La aplicación de fertilizantes nitrogenados y fosfatados al suelo permite suplir gran parte de las necesidades nutricionales del cultivo del arroz, incrementando considerablemente los rendimientos (1). Sin embargo, el empleo irracional de estos productos impacta negativamente los diferentes ecosistemas y encarece el proceso productivo (2). El aislamiento y caracterización de bacterias que se asocian naturalmente al cultivo del arroz y que potencian su crecimiento, ha permitido la elaboración de biopreparados inocuos para el medio ambiente y que provocan incrementos en los rendimientos agrícolas, con la consiguiente disminución de los costos del proceso productivo (3).
Azospirillum y Herbaspirillum se encuentran dentro de los géneros bacterianos más representativos en la rizosfera de las plantas de arroz, aunque también se destacan Bacillus, Pseudomonas, Azotobacter y Burkholderia (4). Sin embargo, los rizobios, bacterias que tradicionalmente se han estudiado por su capacidad de establecer simbiosis con las plantas leguminosas a partir de la formación de nódulos radicales donde realizan la Fijación Biológica de Nitrógeno (FBN) (5), también se han observado colonizando las raíces de las plantas de arroz y como endófitos de este cultivo (6). La inoculación de cepas de rizobios en diversas cultivares de arroz, ha mostrado efectos positivos en el desarrollo, crecimiento y en los rendimientos del cultivo (7, 8).
A pesar de los antecedentes expuestos anteriormente, en el mundo se han realizado escasos estudios sobre la interacción rizobio-arroz y en Cuba hasta el momento, no existen evidencias documentadas donde se demuestre la presencia de los rizobios asociados al cultivo del arroz. Por tal motivo, el objetivo del presente trabajo fue aislar rizobios de la rizosfera de plantas de arroz (Oryza sativa L.) cultivar INCA LP-5, cultivadas en suelo Gley Nodular Ferruginoso Petroférrico. Contar con un cepario de rizobios obtenidos de la rizosfera de plantas de arroz permitiría estudiar detenidamente las particularidades que rigen esta asociación. Con los aislados de rizobios más promisorios se pudieran elaborar biopreparados para incrementar el crecimiento y los rendimientos del arroz, en las condiciones edafoclimáticas de Cuba.
MATERIALES Y MÉTODOS
Muestreo y caracterización química del suelo
Se colectaron plantas de arroz del cultivar INCA LP-5 cultivadas en las terrazas de la zona tres, casillas C y H de la Unidad Científica Tecnológica de Base “Los Palacios” (UCTB-LP), ubicada en la provincia Pinar del Río. Se muestrearon cinco sitios, distantes uno de otro en diagonal, para lograr una representatividad de la microbiota presente en el suelo de la localidad estudiada.
Se tomaron cinco muestras de suelo en cada uno de los sitios muestreados, a una profundidad menor de 30 cm y a más de 20 cm de las raíces de las plantas. Para la determinación del contenido de sodio (Na+), potasio (K+), calcio (Ca2+) y magnesio (Mg2+). Primeramente se extrajeron empleando la técnica de Maslova; el contenido de Na+ y K+, se determinó por fotometría de llama y el de Ca2+ y el Mg2+ empleando EDTA. El contenido de fósforo (P) se evaluó mediante la técnica de Oniani (9). Se determinó además el pH (en KCl) mediante la técnica de potenciometría; así como el contenido porcentual de materia orgánica por el método de Walkley-Black (10).
Aislamiento e identificación de la presencia de rizobios en la rizosfera y rizoplano de las plantas de arroz
En el momento del muestreo, el cultivo se encontraba en la fase vegetativa del crecimiento y sometido a condiciones de inundación, fertilización y riego según lo establecido en el Instructivo Técnico del Arroz (1) y no existían antecedentes de la aplicación de inoculantes a base de rizobios. Se colectaron cuatro plantas de arroz en cada uno de los cinco sitios donde se realizó el muestreo de suelo, para un total de 20 plantas. Se extrajeron las plantas completas, cuidando no dañar el sistema radical y se conservaron a 4°C en bolsas de polietileno para su posterior procesamiento en el laboratorio.
Para el aislamiento de rizobios a partir de suelo rizosférico, se tomaron 10 g de raíces con suelo rizosférico y se colocaron en Erlenmeyer de 250 mL que contenían 100 mL de agua destilada estéril. Los Erlenmeyers se mantuvieron en condiciones de agitación durante 10 min a 60 r min-1. Posteriormente las raíces se extrajeron y a partir de la suspensión de suelo rizosférico obtenida se realizaron diluciones seriadas en agua destilada estéril (10-1-10-6). El aislamiento de los rizobios se realizó mediante la siembra por diseminación de 0,1 mL de las diluciones en placas con medio levadura-manitol-agar (LMA) (11) con rojo congo, a pH 6,8. Las placas se incubaron a 29+1°C durante diez días (12).
Para el aislamiento a partir del rizoplano, se tomaron las raíces extraídas en el procedimiento anterior y se colocaron nuevamente en frascos Erlenmeyer de 250 mL que contenían 100 mL de agua destilada estéril, los cuales se mantuvieron en condiciones de agitación durante otros 30 min a 150 r min-1. Las raíces se retiraron nuevamente y se aplicó a la suspensión obtenida el mismo procedimiento de aislamiento descrita para el suelo rizosférico. Paralelamente, las raíces se cortaron en porciones de 1-2 cm con hojas de bisturí estériles, y posteriormente se colocaron sobre placas que contenían medio Luria-Bertani (13), con el objetivo de permitir el crecimiento de microorganismos más íntimamente adheridos a la superficie radical. Las placas, en ambos casos, se incubaron a 29+1°C durante diez días (14).
Para la purificación de los posibles aislados de rizobios, se tomaron aquellas colonias que presentaron características culturales similares a las descritas para este grupo microbiano (11). Se realizaron pases sucesivos en medio LMA hasta obtener cultivos puros. Los aislados se conservaron a 4°C en tubos de ensayo que contenían dicho medio de cultivo.
Para la selección de rizobios del conjunto de aislados bacterianos obtenidos de la rizosfera de las plantas de arroz cultivar INCA LP-5, se realizó una caracterización cultural y morfológica. Además se tuvo en cuenta el resultado de la prueba de la cetolactasa. Se determinó la capacidad de los aislados para producir ácido/base y se realizaron ensayos de inoculación in vitro en plantas de Macroptillium atropurpureum (DC.) Urb. (siratro).
Caracterización cultural y morfológica de los aislados bacterianos
La pureza de los cultivos bacterianos, así como sus características tintoriales y la morfología de las células se observó mediante la Tinción de Gram (15). Las características morfológicas celulares que se estudiaron fueron, forma de la célula bacteriana, respuesta a la tinción de Gram y la presencia de esporas.
Para la determinación de las características culturales, los aislados se cultivaron por agotamiento en medio LMA con rojo congo, a pH 6,8. Se incubaron durante diez días a 29+1°C y se determinó, mediante microscopía estereoscópica: color, producción de mucus y diámetro (mm) de las colonias (16). Además se determinó la tasa de crecimiento de cada aislado, monitoreando la aparición o no de las colonias en las placas cada 24 h durante diez días de incubación. Los aislados cuyas colonias se hicieran visibles en el medio a los 2-3 días de incubación, se consideraron de rápido crecimiento; las que aparecían a partir de los 4-5 días, se consideraron de crecimiento moderado y aquellas que se observaban a partir de los siete días, se clasificaron como microorganismos de crecimiento lento (11).
Ensayo de la cetolactasa
Los aislados se cultivaron en medio Levadura-Lactosa-Agar (LLA) (17) e incubados a 29+1°C durante tres, cinco y siete días, de acuerdo con su tasa de crecimiento. Posteriormente, se añadieron 10 mL del reactivo de Benedict sobre el crecimiento bacteriano y se mantuvo por 10 min, para observar posible cambio de coloración del medio.
Producción de ácido y base
Los aislados bacterianos se cultivaron en medio LMA a pH 6,8 con indicador azul de bromotimol. A los siete días de incubación a 29+1°C se observó cambio de coloración en el medio de cultivo durante el establecimiento de las colonias bacterianas sobre el mismo.
Ensayo de inoculación in vitro en plantas de siratro
Se prepararon inoculantes de cada uno de los aislados bacterianos, para lo cual se utilizaron Erlenmeyers de 20 mL con 5 mL de medio líquido LM (levadura manitol) a pH 6,8. Se inocularon con una asada de los microorganismos conservados a 4°C en medio sólido. Los frascos se mantuvieron en condiciones de agitación en zaranda orbital a 150 r min-1 durante 18-20 h a 29+1°C.
La identidad de los rizobios se confirmó mediante su capacidad de nodular la leguminosa siratro, tradicionalmente utilizada como planta modelo para este tipo de estudios (18). Las semillas de esta leguminosa se donaron por la Estación Experimental de Pastos y Forrajes de Cascajal, provincia Villa Clara.
Para la descarificación de las semillas se mantuvieron en etanol al 70 % durante cinco minutos, posteriormente se lavaron con agua destilada y se sometieron a un tratamiento con ácido sulfúrico concentrado durante 10 min. Luego se sumergieron en hipoclorito de sodio al 25 % (v/v) durante 15 minutos y se lavaron seis veces con agua destilada estéril. Seguidamente, las semillas se colocaron en placas con Agar-Agua (0,75 g de agar en 100 mL de agua destilada) e incubadas a 29+1°C en la oscuridad durante 24 h. Las semillas germinadas con raíces de aproximadamente 1-2 cm de largo se colocaron en frascos de 200 mL que contenían 50 mL de medio Norris y Date semisólido (15), a razón de una semilla por frasco. Posteriormente, las semillas se inocularon con 0,5 mL de inóculo de los diferentes aislados a una concentración de 108 UFC mL-1.
Las plantas crecieron en condiciones controladas, con un fotoperíodo de 12 h luz/12 h oscuridad, a una temperatura día/noche de 26/22°C y humedad relativa del 70 %. Cinco semanas después, se determinó el número de nódulos en la raíz principal; número total de nódulos; la efectividad de los nódulos y la efectividad de la nodulación (EN) según la expresión:
donde: NNt y NNte son los valores absolutos del número de nódulos totales y número de nódulos totales efectivos, respectivamente. Se determinó, además, la masa seca de las raíces de las plantas (g).
La efectividad de los nódulos se determinó mediante el método visual, lo cual se realizó mediante la disección de los nódulos con hojas de bisturí de acero inoxidable, detectando o no la presencia de una coloración rojiza en el interior de los nódulos, característico de la proteína leghemoglobina (19).
Se empleó un diseño completamente aleatorizado, utilizando siete repeticiones por tratamiento. Se utilizó un tratamiento control, sin inocular. Los resultados procedentes de las variables: número de nódulos en la raíz principal y total, así como el número de nódulos efectivos en cada caso, fueron sometidos a análisis no paramétricos de Kruskal Wallis. En caso de determinar diferencias significativas entre tratamientos, se realizó la prueba de Mann Witney para establecer diferencias entre los rangos promedios de las variables en cada uno de los tratamientos establecidos (20). La variable EN no se sometió a análisis estadístico pues es resultante de los valores absolutos de la nodulación efectiva total y la nodulación total en cada uno de los tratamientos.
Los datos procedentes de la masa seca radical de las plantas de siratro se sometieron a la prueba de normalidad (test de Bartlett) y homogeneidad de varianza (test de Kormogorov-Smirnov) (21). Posteriormente se aplicó análisis de varianza de clasificación simple, utilizando el test de comparación de medias de Tukey para p<0,05 (22); con el objetivo de determinar diferencias entre las medias (20). Estos datos se graficaron con el programa SigmaPlot 2001.
RESULTADOS Y DISCUSIÓN
El suelo Gley Nodular Ferruginoso Petroférrico de la UCTB Los Palacios, Pinar del Rio, se caracterizó por presentar un pH cercano a la neutralidad. El contenido de materia orgánica y los niveles de fósforo disponibles resultaron ser altos. Sin embargo, según el Manual de la Dirección General de Suelos y Fertilizantes (23), este suelo presenta un bajo contenido de calcio (Ca2+), magnesio (Mg2+), sodio (Na+) y potasio (K+) (Tabla I).
El valor de pH en el suelo (6,87), propicia el establecimiento de los rizobios, los cuales requieren valores muy similares para su multiplicación (24). Aunque no es determinante, el establecimiento del cultivo del arroz resulta óptimo a pH cercano a la neutralidad (1). El pH del suelo de esta localidad afectaría positivamente el establecimiento del cultivo y de los rizobios, así como la interacción entre ambos organismos.
Los contenidos de calcio, magnesio, sodio y potasio en este suelo son bajos, lo que pudiera afectar negativamente la multiplicación de los rizobios. Sin embargo, los altos contenidos de fósforo y de materia orgánica que posee, propician la proliferación de estas bacterias y el establecimiento y desarrollo de las plantas de arroz. La materia orgánica condiciona, en gran medida, la distribución de los principales grupos microbianos en el suelo y potencia su crecimiento en aquellas zonas donde existe una mayor disponibilidad de los nutrientes (25).
En el momento del muestreo, las plantas de arroz cultivar INCA LP-5 se encontraban en condiciones de inundación, propiciada por el riego al que estuvo sometido el cultivo y por las características descritas con anterioridad para este tipo de suelo. Se plantea que el suelo de la UCTB Los Palacios clasifica como Hidromórfico Gley Nodular Ferruginoso Petroférrico, de textura fina, con un gran contenido de arcillas y de poco drenaje (26).
Una relación directamente proporcional entre el contenido de humedad del suelo y la distancia recorrida por Azospirillum se describió con anterioridad (27). Esto sugiere que la elevada humedad presente en el suelo muestreado a las que estaban sometidas las plantas de arroz cultivar INCA LP-5, pudiera haber favorecido el movimiento de bacterias flageladas como los rizobios hacia las raíces.
El proceso de aislamiento de rizobios de la rizosfera de las plantas de arroz cultivar INCA LP-5 se realizó tomando como criterio primario las características culturales de las colonias descritas para el grupo de los rizobios (15). En el medio LMA, las cepas de crecimiento rápido originan colonias mucosas de 1 a 5 mm luego de 3 a 5 días de incubación, mientras que en las de crecimiento lento, con una superficie más seca, las colonias no exceden un diámetro de 1 mm en un período de diez días (15).
Para la selección de las colonias típicas de rizobios, otro elemento esencial fue la falta de absorción del rojo congo, pues generalmente los cultivos jóvenes de estos microorganismos forman colonias translúcidas, blanquecinas o ligeramente rosadas en el centro (28). Teniendo en cuenta lo anterior, se aislaron 78 colonias que reunían las características de tamaño y mucosidad típicas de los rizobios, pues se observaron como colonias grandes (2-4 mm) y mucosas, a los 2-3 días; excepto un aislado en el cual se visualizaron colonias secas de 1-2 mm de diámetro a los 4-5 días de incubación.
El 44,87 % de las 78 colonias bacterianas se obtuvieron a partir de las muestras de suelo rizosférico, mientras que el 55,13 % restante, se encontraron en el rizoplano de las plantas de arroz. De este último grupo, el 90 % de los aislados se detectaron ligeramente adheridos al rizoplano, ya que se obtuvieron en las diluciones seriadas que se realizaron con las raíces de las plantas. El 10 % restante, se identificó sobre el tejido epitelial, observándose como una película fina, mucosa y blanquecina que cubría las raíces a los cinco días de haberlas colocado sobre el medio Luria-Bertani.
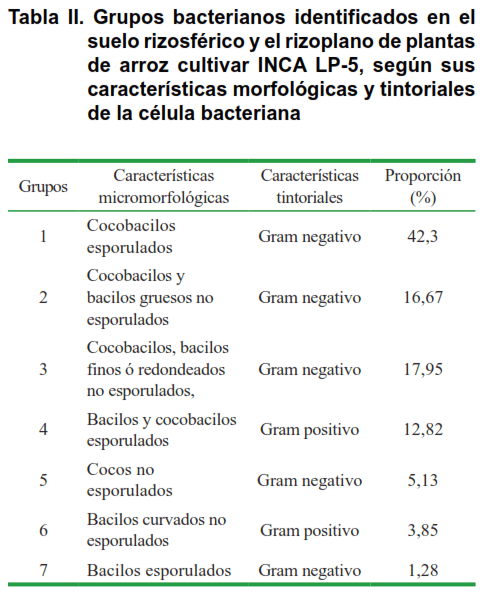
La tinción de Gram se empleó como segundo criterio de clasificación. Los resultados de esta determinación permitieron agrupar a los 78 aislados en siete grupos, considerando las características morfológicas de las células bacterianas, así como la respuesta a la Tinción de Gram (Tabla II).
La caracterización morfológica y tintorial permitió identificar tres tipos morfológicos de células: uno cocobacilar (predominante) y otros dos donde las células presentaban forma de cocos o bacilos. La mayor parte de los aislados rizosféricos (42,3 %) correspondieron a cocobacilos esporulados, Gram negativos; mientras que el grupo menos representativo (1,28 %) estuvo conformado por bacilos esporulados Gram negativos.
La tinción de Gram permitió observar una gran diversidad en las células bacterianas en la rizosfera de las plantas de arroz cultivar INCA LP-5. El alto contenido de materia orgánica y de fósforo presente en el suelo de las áreas arroceras de la UCTB Los Palacios, pudiera estar favoreciendo el establecimiento de la diversidad bacteriana que se observó en esta investigación.
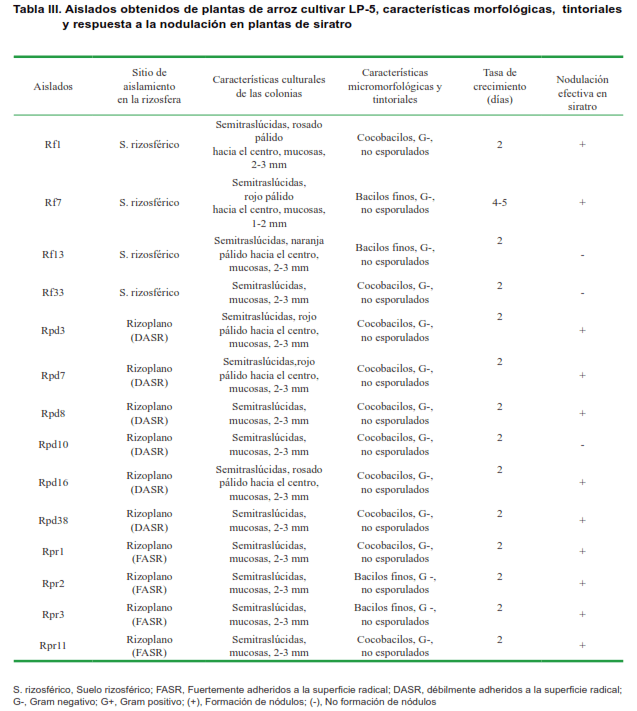
Los rizobios se han descrito como bacilos finos, Gram negativos, no esporulados (17). Estas características son similares a las que se observaron en el 17,95 % de los aislados bacterianos (Tabla II, grupo 3); ya que se observaron como cocobacilos o bacilos finos, con extremos redondeados y en ocasiones afinados, no esporulados y con respuesta negativa a la tinción de Gram. Este porcentaje corresponde con catorce aislados, de los cuales el 21,43 % (cuatro aislados) se obtuvieron del suelo rizosférico y el 78,57 % (once aislados) se obtuvieron a partir del rizoplano (Tabla III).
Todos los aislados que se estudiaron produjeron colonias semitranslúcidas en el medio LMA (pH 6,8); sin embargo, se observó una diversidad de coloraciones tenues hacia el centro de las colonias, sobre todo en aquellos aislados provenientes del suelo rizosférico de las plantas de arroz. Resultados alcanzados por otros autores difieren a los obtenidos en esta investigación, ya que obtuvieron colonias blanquecinas en el 100 % de los aislados de rizobios que se estudiaron (29).
Se realizó la prueba de la cetolactasa, con el objetivo de diferenciar los aislados en estudio del género Agrobacterium ya que ambos grupos presentan características culturales y morfológicas similares. Por otra parte, el género Agrobacterium se estudia usualmente por su capacidad de producir agallas o tumores en las raíces y tallo de las plantas, estructura muy similar a los nódulos que forman los rizobios en las plantas leguminosas (30).
La prueba de la cetolactasa, permite observar la capacidad de Agrobacterium (no en los géneros de la familia Rhizobiaceae) para reducir el óxido de cobre I (Cu2O) presente en el reactivo de Bénedict que se emplea en esta prueba (31). En este trabajo todos los aislados seleccionados como posibles rizobios, por sus características culturales y morfológicas, resultaron cetolactasa negativos. Esta respuesta corrobora la posible ubicación taxonómica de los aislados en estudio como rizobios (32). Al determinar la capacidad de producir ácido o base en medio LMA con azul de bromotimol, se observó en todos los casos un cambio de coloración de verde a amarillo alrededor del crecimiento bacteriano, lo cual indica la capacidad de estos microorganismos para producir ácido.
Los ácidos orgánicos que se producen por los microorganismos rizosféricos incrementan la disponibilidad de micronutrientes como el fósforo, el hierro, el zinc y el manganeso en el suelo, al disminuir el pH en la rizosfera, o por la quelación de estos micronutrientes. La detoxificación de metales en el suelo se ha informado como otra de las funciones que realizan los ácidos orgánicos que escretan por estos microorganismos (33).
Los estudios de la producción de ácidos orgánicos por los rizobios mayormente se enfocan a la promoción del crecimiento vegetal, como por ejemplo el ácido 3-indolacético (AIA) y aquellos que se relacionan con la solubilización de fosfatos en los suelos (34, 35).
La tasa de crecimiento de los rizobios es uno de los aspectos a tener en cuenta durante los estudios taxonómicos, ya que su capacidad de multiplicarse rápida, moderada o lentamente en el medio de cultivo, es una característica fenotípica muy relacionada al género al cual pertenecen las especies de este grupo bacteriano (28).
El estudio de la tasa de crecimiento de los diferentes aislados sobre el medio LMA a pH 6,8 permitió detectar un crecimiento rápido en la mayoría de los aislados, ya que pudo constatarse al menos una colonia de estos microorganismos sobre el medio de cultivo a las 48 horas de incubación. El aislado Rf7, a diferencia del resto, presentó un crecimiento moderado pues sus colonias se comenzaron a visualizar a partir del cuarto-quinto día de incubación (Tabla III).
Sólo el 8 al 20 % de la superficie de las raíces se coloniza por las bacterias. En las zonas de mayor exudación se observa un mayor establecimiento microbiano (36). Una mayor efectividad en la colonización de la rizosfera por ciertos microorganismos se debe, entre otros factores, a su rápida multiplicación. Esta capacidad les permite utilizar una gran cantidad y diversidad de nutrientes presentes en los exudados radicales como aminoácidos, ácidos orgánicos, azúcares, vitaminas, enzimas, iones inorgánicos y moléculas gaseosas como fuentes de carbono, oxígeno, nitrógeno, fósforo y azufre (37).
La capacidad de producir nódulos como parte de la simbiosis rizobio-leguminosa, constituye una de las premisas a tener en cuenta para la ubicación taxonómica de una rizobacteria en el grupo de los rizobios (8). Siratro constituye una planta modelo para evaluar la capacidad de infección y la efectividad de la nodulación de aislados de bacterias bajo condiciones controladas. Esta planta se utiliza en investigaciones de este tipo, pues responde como hospedero a un amplio espectro de cepas de rizobios provenientes de una gran diversidad de leguminosas (38).
El ensayo de inoculación in vitro en las plantas de siratro permitió observar que todos, excepto Rf13, Rf33 y Rpd10, produjeron nódulos efectivos en las raíces de estas plantas (Tabla III). El ensayo permitió concluir que 11 aislados bacterianos provenientes del suelo rizosférico y del rizoplano de plantas de arroz INCA LP-5, pertenecen al grupo de los rizobios. Estos resultados constituyen la primera evidencia en Cuba de la interacción de bacterias del grupo de los rizobios con el cultivo del arroz.
Los resultados que se obtuvieron en esta investigación fueron similares a los que se encontraron en los primeros estudios sobre la interacción de los rizobios con el cultivo del arroz (39). En estas investigaciones se demostró, a través de bioensayos con las leguminosas trébol blanco (Trifolium repens var. Dutch) y alfalfa (Medicago sativa var. Gemini), que eran rizobios los endófitos que se encontraron en las plantas de arroz. Estos microorganismos produjeron nódulos en dichas leguminosas.
Los avances en técnicas moleculares han permitido realizar una reubicación genérica de especies bacterianas dentro del grupo de los rizobios, teniendo en cuenta su capacidad de realizar la FBN y de formar nódulos radiculares en las plantas leguminosas. Actualmente 14 géneros bacterianos conforman al grupo de los rizobios,de ellos 11 son Rhizobium, Mesorhizobium, Ensifer, Bradyrhizobium, Phyllobacterium, Microvirga, Azorhizobium, Ochrobactrum, Methylobacterium, Devosia y Shinella, en la clase alfa-proteobacterias; dos: Burkholderia y Cupriavidus, dentro de la clase beta-proteobacterias y el género Pseudomonas en la clase gamma-proteobacterias (5).
A pesar de que la caracterización que se realizó en es te trabajo no es suficiente para ubicar taxonómicamente a los aislados de rizobios, en todos ellos; con excepción de Rf7, se identificaron aspectos fenotípicos de la familia Rhizobiaceae (40). La formación de nódulos efectivos en la FBN, la produción de ácido y rápido crecimiento en medio LMA, permitiría clasificarlos dentro de los géneros Rhizobium, Ensifer o Shinella, de la familia Rhizobiaceae.
El aislado Rf7, que también induce la formación de nódulos, produce ácido y presenta una tasa de crecimiento moderada pudiera ubicarse dentro del género Mesorhizobium, en la familia Phyllobacteriaceae (41, 42).
La determinación de la presencia de 11 aislados de rizobios en la rizosfera de las plantas de arroz cultivar INCA LP-5, resultados que se muestran en esta investigación, constituye la primera evidencia documentada en Cuba sobre la interacción de rizobio-arroz. La implementación de estos resultados en la investigación básica y en la elaboración de bioproductos para el cultivo del arroz, abre nuevas puertas al conocimiento de esta interacción en el país.
El aislamiento de rizobios de la rizosfera de plantas de arroz cultivar INCA LP-5 que se realizó en este trabajo, permitió identificar una mayor cantidad y diversidad de estos microorganismos en el rizoplano que en el suelo rizosférico de estas plantas (Tabla III). El rizoplano es la zona de la rizosfera más influenciada por los exudados radicales y donde la concentración de estos compuestos es máxima, pues una vez en el suelo, los exudados se van dispersando (43). La presencia de altas concentraciones de nutrientes en el rizoplano, lo convierte en una zona atractiva para la colonización microbiana. Esto pudiera explicar el qie se encontrara mayor número y diversidad de rizobios en el rizoplano respecto al suelo risoférico.
Los exudados radicales establecen un ambiente selectivo en la rizosfera, de manera que propician la colonización de ciertos microorganismos y desfavorecen el desarrollo de otros (38). Diferencias en los requerimientos nutricionales de los microorganismos en la rizosfera, así como sus respuestas quimiotácticas, cobran vital importancia en la especificidad entre las plantas y los microorganismos (44).
Diferencias en la movilidad de cada aislado de rizobio, su respuesta quimiotáctica a los exudados radicales, el efecto selectivo que ejerce la especificidad de estos compuestos, así como las características genotípicas y fisiológicas intrínsecas del cultivar de arroz INCA LP-5, pudieran haber influido en la localización de los rizobios en la rizosfera de las plantas estudiadas.
En esta investigación además, se pudo constatar diferencias entre los aislados de rizobios que se obtuvieron del rizoplano de las plantas, teniendo en cuenta su capacidad para adherirse a la superficie radical. Un grupo de estos aislados se caracterizó por adherirse más estrechamente a las células epidérmicas de la raíz y otro mantuvo una relación menos estrecha con la superficie radical (Tabla III). Cuatro de los aislados de rizobios (Rpr1, Rpr2, Rpr3 y Rpr11) se encontraron fuertemente unidos a la superficie radical. Se ha documentado la capacidad de los rizobios para la formación de biopelículas sobre el rizoplano, mecanismo mediante el cual logra mantener una estrecha relación con los exudados de las raíces de las plantas (45).
Estudios genómicos han aportado elementos importantes que explican los mecanismos involucrados en la expresión de los genes de RapA1. Esta proteína tiene una importancia crucial en la unión de los rizobios al rizoplano y en la formación de las biopelículas en las raíces de las plantas (39). La presencia de flagelos en los rizobios, así como la producción de exopolisacáridos, también tienen una función importante en la formación de estas asociaciones, ya que mutaciones en genes relacionados con la síntesis de estos componentes afectan la estabilidad de las biopelículas (40).
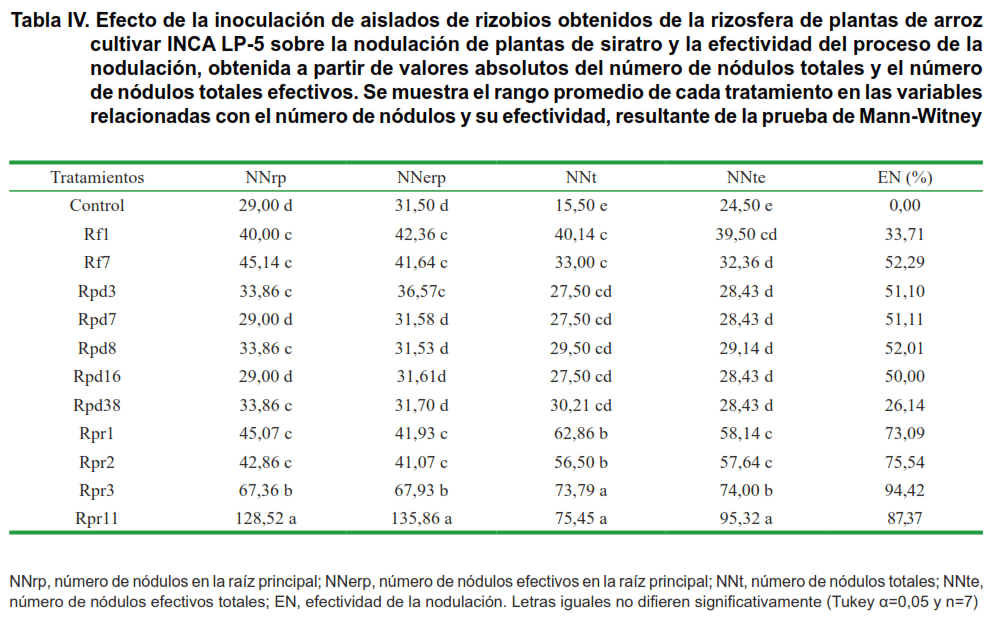
El efecto de la inoculación de los 11 aislados que produjeron nódulos efectivos en las plantas de siratro y que se consideran rizobios se muestra en la Tabla IV.
En las plantas que se inocularon con el aislado Rpr11 se observaron los mejores resultados en el número de nódulos. Se observaron diferencias significativas entre Rpr11 y el resto de los aislados, excepto con Rpr3 en el número de nódulos totales. Ambos aislados, además, mostraron los mayores porcentajes de EN.
Además de los aislados mencionados anteriormente, se destacaron Rpr1 y Rpr2, que superaron al resto en las variables número de nódulos totales, número de nódulos totales efectivos y en la efectividad del proceso.
La abundante nodulación en las plantas de siratro que se inocularon con Rpr1, Rpr2, Rpr3 y Rpr11,microorganismos que se encontraron fuertemente adheridos a la superficie radical, permite sugerir la presencia de mecanismos en estos microorganismos que les permite colonizar más efectivamente diferentes puntos de la raíz e invadir los pelos radicales de las plantas de siratro. Ambas fases constituyen las primeras etapas de la formación de los nódulos (46, 47).
Los nódulos que se ubican sobre la raíz principal poseen una mayor efectividad en el aporte de nitrógeno fijado, lo cual se traduce en un incremento en el desarrollo foliar y los rendimientos del cultivo. La cercanía de estas estructuras a los vasos conductores hace que el microsimbionte tenga un mayor acceso a los fotosintatos, y la planta al nitrógeno proveniente de la FBN que realizan los rizobios (48).
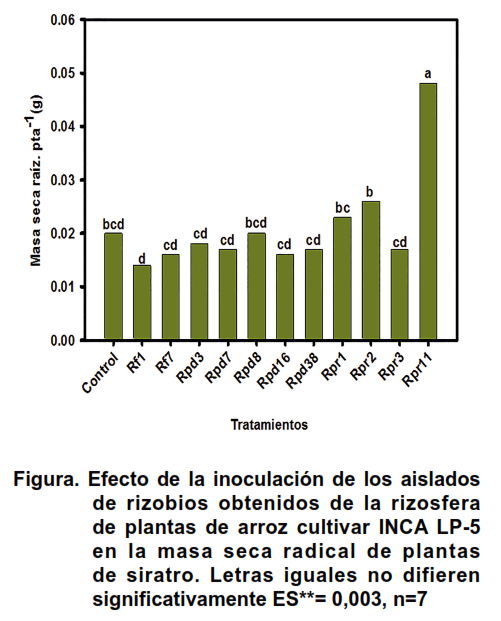
El amonio que se produce durante la fijación del nitrógeno atmosférico por las células de rizobios se emplea por la leguminosa para la síntesis de aminoácidos, enzimas y proteínas estructurales (49). La disección de los nódulos presentes en las raíces de las plantas de siratro permitió observar que todos los aislados bacterianos produjeron nódulos efectivos, ya que se observó una coloración rojiza en el interior de estas estructuras, característica de la presencia de la leghemoglobina. Esta proteína constituye un indicador de la efectividad de los nódulos para realizar el proceso de fijación de nitrógeno y del establecimiento exitoso de la simbiosis rizobio-leguminosa (20). La presencia de un gran número de nódulos en la raíz principal de las plantas de siratro inoculadas con Rpr11, muchos de los cuales fueron efectivos en la FBN, pudiera explicar el efecto positivo de este aislado sobre la masa seca de las raíces de las plantas (Figura).
El número y efectividad de los nódulos de las plantas que se inocularon con Rpr3 superó a aquellas donde se empleó Rpr2. Sin embargo, este microorganismo produjo un mayor efecto en el crecimiento radical. El comportamiento pudiera deberse a una mayor actividad de la enzima nitrogenasa en Rpr2. Este aislado produciría entonces mayores niveles de amonio que Rpr3, traduciéndose en un incremento del crecimiento radical de las plantas de siratro. Resultados similares se encontraron en soya, pues se observaron incrementos de la altura de las plantas ante un aumento de la actividad de la enzima nitrogenasa (50).
Los aislados que produjeron el mayor número de nódulos en las raíces de las plantas (Rpr1, Rpr2, Rpr3 y Rpr11) coincidieron con aquellos que presentaron los porcentajes de EN más elevados, y en el caso de Rpr11 también con un efecto superior en la masa seca radical. Con este ensayo se confirma la pertenencia de estos aislados al grupo de los rizobios y se destacan algunos en su capacidad de nodular plantas de siratro.
CONCLUSIONES
Las características del suelo Gley Nodular Ferruginoso Petroférrico de la UCTB Los Palacios, Pinar del Río propiciaron la proliferación de poblaciones de rizobios en la rizosfera de plantas de arroz cultivar INCA LP-5. Esta investigación permitió contar con nuevos aislados de rizobios que defirieron en el nivel de interacción que establecieron en la rizosfera de las plantas de arroz y en su efectividad en la nodulación de siratro. Estos hallazgo constituyen la primera evidencia en Cuba donde se aborda la interacción de los rizobios con el cultivo del arroz.
NOTAS AL PIE
AConsejo Internacional de cereales. Informe de Mercado de Cereales. Resumen de las perspectivas para cereales y oleaginosas clave, Inst. Consejo Internacional de cereales, 2015, 8 p.
BFAO y IAEA. Programme. Rice production: what Cuba can teach the world? Nuclear Techniques in Food and Agriculture. Inst. FAO, 2014, Austria, 1 p.
BIBLIOGRAFÍA
1. MINAGRI. Instructivo Técnico del Arroz. Ed. Centro Nacional de Sanidad Vegetal, Instituto de Investigaciones del Arroz, 2005, Cuba, 22 p., ISBN 959-246-037-X.
2. Biau, A.; Santiveri, F.; Mijangos, I. y Lloveras, J. “The impact of organic and mineral fertilizers on soil quality parameters and the productivity of irrigated maize crops in semiarid regions”. European Journal of Soil Biology, vol. 53, 2012, pp. 56-61, ISSN 1164-5563, DOI 10.1016/j.ejsobi.2012.08.008.
3. Mehnaz, S. “Azospirillum: A Biofertilizer for Every Crop” [en línea]. En: ed. Kumar N. A., Plant Microbes Symbiosis: Applied Facets, Ed. Springer India, New Delhi, 2015, pp. 297-314, ISBN 978-81-322-2067-1, [Consultado: 16 de noviembre de 2016], Disponible en: <http://link.springer.com/10.1007/978-81-322-2068-8_15>.
4. Hernández, R. A.; Rives, R. N.; Acebo, G. Y.; Díaz, de la O. A.; Heydrich, P. M. y Divan, B. V. L. “Potencialidades de las bacterias diazotróficas asociativas en la promoción del crecimiento vegetal y el control de Pyricularia oryzae (Sacc.) en el cultivo del arroz (Oryza sativa L.)”. Revista de Protección Vegetal, vol. 29, no. 1, 2014, pp. 1-10, ISSN 1010-2752.
5. Berrada, H. y Fikri-Benbrahim, K. “Taxonomy of the Rhizobia: Current Perspectives”. British Microbiology Research Journal, vol. 4, no. 6, 2014, pp. 616-639, ISSN 2231-0886, DOI 10.9734/BMRJ/2014/5635.
6. Chen, C. y Zhu, H. “Are common symbiosis genes required for endophytic rice-rhizobial interactions?”. Plant Signaling & Behavior, vol. 8, no. 9, 2013, ISSN 1559-2324, DOI 10.4161/psb.25453, [Consultado: 16 de noviembre de 2016], Disponible en: <http://www.tandfonline.com/doi/abs/10.4161/psb.25453>.
7. Mia, M. A. B.; Shamsuddin, Z. H. y Mahmood, M. “Effects of rhizobia and plant growth promoting bacteria inoculation on germination and seedling vigor of lowland rice”. African Journal of Biotechnology, vol. 11, no. 16, 2012, pp. 3758–3765, ISSN 1684-5315, DOI 10.5897/AJB09.1337.
8. Hasan, M.; Bano, A.; Hassan, S. G.; Iqbal, J.; Awan, U.; Rong-ji, D. y Khan, K. A. “Enhancement of rice growth and production of growth-promoting phytohormones by inoculation with Rhizobium and other rhizobacteria”. World Applied Sciences Journal, vol. 31, no. 10, 2014, pp. 1734-1743, ISSN 1818-4952, DOI 10.5829/idosi.wasj.2014.31.10.364.
9. Oniani, O. G. “Determinación del fósforo y potasio del suelo en una misma solución de los suelos Krasnozen y Podsólicos en Georgia”. Agrojima, vol. 6, 1964, p. 25.
10. Walkley, A. y Black, I. A. “An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method.”Soil Science, vol. 37, no. 1, 1934, pp. 29–38, ISSN 0038-075X, 1538-9243.
11. Wang, T.; Romero-Martínez, J. y López-Lara, I. “Rhizobium y su destacada simbiosis con plantas”. En: Microbiología en línea, Ed. Universidad Nacional Autónoma de México, México, 2001, pp. 344-386, ISBN 978-968-36-8879-8.
12. Vincent, J. M. A manual for the practical study of root-nodule bacteria. (ser. IBP Handbook, no. ser. 15), Ed. International Biological Programme, 1970, Oxford, 164 p., ISBN 978-0-632-06410-6.
13. Bertani, G. “Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli”. Journal of Bacteriology, vol. 62, no. 3, 1951, pp. 293-300, ISSN 0021-9193.
14. Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; von Mering, C. y Vorholt, J. A. “Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice”. The ISME Journal, vol. 6, no. 7, 2012, pp. 1378-1390, ISSN 1751-7362, DOI 10.1038/ismej.2011.192.
15. Norris, D. O. y Date, R. A. “Legume bacteriology”. En: Shaw N. H. y Bryan W. W., Tropical pasture research: principles and methods, (ser. Commonwealth Bureau of Pastures and Field Crops, no. ser. 51), Ed. Commonwealth Agricultural Bureaux, Farnham Royal, England, 1976, pp. 134-174, ISBN 978-0-85198-358-5.
16. Frioni, L. “Ecología microbiana del suelo”. En: Procesos Microbianos, Ed. Editorial de la fundación universidad nacional de rio cuarto, Montevideo, 1990, pp. 205–253, ISBN 950-665-110.
17. Thies, J. E.; Singleton, P. W. y Bohlool, B. B. “Modeling Symbiotic Performance of Introduced Rhizobia in the Field by Use of Indices of Indigenous Population Size and Nitrogen Status of the Soil”. Applied and Environmental Microbiology, vol. 57, no. 1, 1991, pp. 29-37, ISSN 0099-2240, 1098-5336.
18. López-López, A.; Rogel-Hernández, M. A.; Barois, I.; Ortiz Ceballos, A. I.; Martínez, J.; Ormeno-Orrillo, E. y Martínez-Romero, E. “Rhizobium grahamii sp. nov., from nodules of Dalea leporina, Leucaena leucocephala and Clitoria ternatea, and Rhizobium mesoamericanum sp. nov., from nodules of Phaseolus vulgaris, siratro, cowpea and Mimosa pudica”. International Journal of Systematic and Evolutionary Microbiology, vol. 62, no. 9, 2012, pp. 2264-2271, ISSN 1466-5026, 1466-5034, DOI 10.1099/ijs.0.033555-0.
19. Vázquez-Limón, C.; Hoogewijs, D.; Vinogradov, S. N. y Arredondo-Peter, R. “The evolution of land plant hemoglobins”. Plant Science, vol. 191-192, 2012, pp. 71-81, ISSN 0168-9452, DOI 10.1016/j.plantsci.2012.04.013.
20. Sigarroa, A. Biometría y Diseño Experimental. 1.a ed., Ed. Pueblo y Educación, 1985, La Habana, Cuba, 393 p.
21. Allen, M. E. Kolmogorov-Smirnov test for discrete distributions [en línea]. Master Thesis, Naval Postgraduate School, marzo de 1976, Monterey, California, 93 p., [Consultado: 26 de mayo de 2016], Disponible en: <http://calhoun.nps.edu/handle/10945/17830>.
22. Tukey, J. W. “Bias and confidence in not quite large samples”. The Annals of Mathematical Statistics, vol. 29, no. 2, junio de 1958, pp. 614-623, ISSN 0003-4851, DOI 10.1214/aoms/1177706647.
23. Mesa, A.; Naranjo, M.; Cancio, R.; Martín, A.; Clemente, B.; Suárez, O. y Pacheco, E. Manual de interpretación de los índices físico-químicos y morfológicos de los suelos cubanos. 2.a
ed., Ed. Pueblo y Educación, 1984, La Habana, Cuba, 10 p.
24. Marquina, M. E.; González, N. E. y Castro, Y. “Caracterización fenotípica y genotípica de doce rizobios aislados de diversas regiones geográficas de Venezuela”. Revista de Biología Tropical, vol. 59, no. 3, 2011, pp. 1017-1036, ISSN 0034-7744.
25. Ruiz, S. “Influencia de microorganismos sobre caracteristicas fisicoquimicos de los suelos de cultivo de cacao (Theobroma cacao L.), en tingo maria”. Revista Investigación y Amazonía, vol. 2, no. 1,2, 2013, pp. 67-74, ISSN 2224-445X.
26. Hernández, A.; Ascanio, M. O.; Morales, M. y León, A. La historia de la clasificación de los suelos de Cuba. Ed. Félix Varela, 2006, La Habana, Cuba, 98 p., ISBN 959-07-0145-0.
27. Bashan, Y.; Holguín, G. y Ferrera-Cerrato, R. “Interacciones entre plantas y microorganismos benéficos. III. Procedimientos para el aislamiento y caracterización de hongos micorrízicos y rizobacterias promotoras del crecimiento de plantas”. Terra, vol. 14, no. 2, 1996, pp. 159-194, ISSN 0187-5779.
28. Durán, D.; Rey, L.; Sánchez-Cañizares, C.; Jorrín, B.; Imperial, J. y Ruiz-Argüeso, T. “Biodiversity of Slow-Growing Rhizobia: The Genus Bradyrhizobium”. En: eds. Rodelas G. M. B. y Gonzalez-López J., Beneficial plant-microbial interactions: ecology and applications, Ed. CRC Press, Boca Raton, FL, 2013, pp. 20-37, ISBN 978-1-4665-8717-5.
29. Mohamed, A. T. H. y Saifelden, A. M. “Diversity of Rhizobium leguminosarum bv. viceae Strains Isolated from Different Schemes in Shendi Area”. Extensive Journal of Applied Sciences, vol. 3, no. 1, 2015, pp. 1-10, ISSN 2409-9511.
30. Peix, A.; Ramírez-Bahena, M. H.; Velázquez, E. y Bedmar, E. J. “Bacterial Associations with Legumes”. Critical Reviews in Plant Sciences, vol. 34, no. 1-3, 2015, pp. 17-42, ISSN 0735-2689, 1549-7836, DOI 10.1080/07352689.2014.897899.
31. Clark, A. G. “A Selective Medium for the Isolation of Agrobacterium Species”. Journal of Applied Bacteriology, vol. 32, no. 3, septiembre de 1969, pp. 348-351, ISSN 0021-8847, DOI 10.1111/j.1365-2672.1969.tb00982.x.
32. Lloret, L. y Martínez-Romero, E. “Evolución y filogenia de Rhizobium”. Revista Latinoamericana de Microbiología, vol. 47, no. 1-2, 2005, pp. 43–60, ISSN 0034-9771.
33. Singh, Y.; Ramteke, P. W. y Shukla, P. K. “Characterization of Rhizobium isolates of pigeon pea rhizosphere from Allahabad soils and their potential pgpr characteristics”. International Journal of Research in Pure and Applied Microbiology, vol. 3, no. 1, 2013, pp. 4–7, ISSN 2277-3843.
34. Panhwar Q. A. “Root colonization and association of phosphate- solubilizing bacteria at various levels of triple supper phosphate in aerobic rice seedlings”. African Journal of Microbiology Research, vol. 6, no. 10, 2012, pp. 2277-2286, ISSN 1996-0808, DOI 10.5897/AJMR11.916.
35. Zúñiga, A.; Poupin, M. J.; Donoso, R.; Ledger, T.; Guiliani, N.; Gutiérrez, R. A. y González, B. “Quorum Sensing and Indole-3-Acetic Acid Degradation Play a Role in Colonization and Plant Growth Promotion of Arabidopsis thaliana by Burkholderia phytofirmans PsJN”. Molecular Plant-Microbe Interactions, vol. 26, no. 5, 2013, pp. 546-553, ISSN 0894-0282, DOI 10.1094/MPMI-10-12-0241-R.
36. de la Fe Pérez, Y.; Díaz de la Osa, A.; Restrepo-Franco, G. M.; Diván-Baldani, V. L. y Hernández-Rodríguez, A. “Diversidad de bacterias diazotróficas asociativas potencialmente eficientes en cultivos de importancia económica”. Revista Cubana de Ciencias Biológicas, vol. 4, no. 1, 2015, pp. 17-26, ISSN 2307-695X.
37.Badri, D. V.; Chaparro, J. M.; Zhang, R.; Shen, Q. y Vivanco, J. M. “Application of Natural Blends of Phytochemicals Derived from the Root Exudates of Arabidopsis to the Soil Reveal That Phenolic-related Compounds Predominantly Modulate the Soil Microbiome”. Journal of Biological Chemistry, vol. 288, no. 7, 2013, pp. 4502-4512, ISSN 0021-9258, 1083-351X, DOI 10.1074/jbc.M112.433300.
38. Lima, A. S.; Nóbrega, R. S. A.; Barberi, A.; da Silva, K.; Ferreira, D. F. y Moreira, F. M. de S. “Nitrogen-fixing bacteria communities occurring in soils under different uses in the Western Amazon Region as indicated by nodulation of siratro (Macroptilium atropurpureum)”. Plant and Soil, vol. 319, no. 1-2, 2009, pp. 127-145, ISSN 0032-079X, 1573-5036, DOI 10.1007/s11104-008-9855-2.
39. Yanni, Y. G.; Rizk, R. Y.; Corich, V.; Squartini, A.; Ninke, K.; Philip-Hollingsworth, S.; Orgambide, G.; de Bruijn, F.; Stoltzfus, J.; Buckley, D.; Schmidt, T. M.; Mateos, P. F.; Ladha, J. K. y Dazzo, F. B. “Natural endophytic association between Rhizobium leguminosarum bv. trifolii and rice roots and assessment of its potential to promote rice growth”. Plant and Soil, vol. 194, no. 1/2, 1997, pp. 99-114, ISSN 0032-079X, DOI 10.1023/A:1004269902246.
40. Santi, C.; Bogusz, D. y Franche, C. “Biological nitrogen fixation in non-legume plants”. Annals of Botany, vol. 111, no. 5, 2013, pp. 743-767, ISSN 0305-7364, 1095-8290, DOI 10.1093/aob/mct048.
41. Degefu, T.; Wolde-meskel, E.; Liu, B.; Cleenwerck, I.; Willems, A. y Frostegard, A. “Mesorhizobium shonense sp. nov., Mesorhizobium hawassense sp. nov. and Mesorhizobium abyssinicae sp. nov., isolated from root nodules of different agroforestry legume trees”. International Journal of Systematic and Evolutionary Microbiology, vol. 63, no. 5, 2013, pp. 1746-1753, ISSN 1466-5026, 1466-5034, DOI 10.1099/ijs.0.044032-0.
42. Zheng, W. T.; Li, Y.; Wang, R.; Sui, X. H.; Zhang, X. X.; Zhang, J. J.; Wang, E. T. y Chen, W. X. “Mesorhizobium qingshengii sp. nov., isolated from effective nodules of Astragalus sinicus”. International Journal of Systematic and Evolutionary Microbiology, vol. 63, no. 6, 2013, pp. 2002-2007, ISSN 1466-5026, 1466-5034, DOI 10.1099/ijs.0.044362-0.
43. de Lima, T. E. F.; Bezerra, J. L. y de Queiroz, C. M. A. “Fungi from the rhizosphere and rhizoplane from the grapevine Vitis labrusca in Pernambuco, Brazil”. Nova Hedwigia, vol. 99, no. 3, 2014, pp. 531-540, ISSN 0029-5035, DOI 10.1127/0029-5035/2014/0204.
44. Philippot, L.; Raaijmakers, J. M.; Lemanceau, P. y van der Putten, W. H. “Going back to the roots: the microbial ecology of the rhizosphere”. Nature Reviews Microbiology, vol. 11, no. 11, 2013, pp. 789-799, ISSN 1740-1526, 1740-1534, DOI 10.1038/nrmicro3109.
45. Nogales, J.; Pérez-Mendoza, D.; Gallegos, M. T. y Soto, M. J. “Importance of Motile and Biofilm Lifestyles of Rhizobia for the Establishment of Symbiosis with Legumes”. En: Rodelas G. M. B. y Gonzalez-López J., Beneficial plant-microbial interactions: ecology and applications, Ed. CRC Press, Boca Raton, FL, 2013, pp. 47-55, ISBN 978-1-4665-8717-5.
46. Frederix, M.; Edwards, A.; Swiderska, A.; Stanger, A.; Karunakaran, R.; Williams, A.; Abbruscato, P.; Sanchez-Contreras, M.; Poole, P. S. y Downie, J. A. “Mutation of praR in Rhizobium leguminosarum enhances root biofilms, improving nodulation competitiveness by increased expression of attachment proteins”. Molecular Microbiology, vol. 93, no. 3, 2014, pp. 464-478, ISSN 0950-382X, DOI 10.1111/mmi.12670.
47. Murray, J. D. “Invasion by Invitation: Rhizobial Infection in Legumes”. Molecular Plant-Microbe Interactions, vol. 24, no. 6, 2011, pp. 631-639, ISSN 0894-0282, DOI 10.1094/MPMI-08-10-0181.
48. McDermott, T. R. y Graham, P. H. “Bradyrhizobium japonicum Inoculant Mobility, Nodule Occupancy, and Acetylene Reduction in the Soybean Root System”. Applied and Environmental Microbiology, vol. 55, no. 10, 1989, pp. 2493-2498, ISSN 0099-2240, 1098-5336.
49. Gil-Quintana, E.; Larrainzar, E.; Seminario, A.; Díaz-Leal, J. L.; Alamillo, J. M.; Pineda, M.; Arrese-Igor, C.; Wienkoop, S. y Gonzalez, E. M. “Local inhibition of nitrogen fixation and nodule metabolism in drought-stressed soybean”. Journal of Experimental Botany, vol. 64, no. 8, 2013, pp. 2171-2182, ISSN 0022-0957, 1460-2431, DOI 10.1093/jxb/ert074.
50. Mubarik, N. R. y Sunatmo, T. I. “Symbiotic of nitrogen fixation between acid aluminium tolerant Bradyrhizobium japonicum and soybean”. En: Ohyama T., Advances in biology and ecology of nitrogen fixation, Ed. Intech, Rijeka, Croatia, 2014, pp. 259-272, ISBN 978-953-51-1216-7.
Recibido: 09/10/2015
Aceptado: 22/04/2016
M.Cs. Ionel Hernández Forte, Instituto Nacional de Ciencias Agrícolas (INCA), Gaveta postal No.1, San José de las Lajas. Mayabeque, Cuba. CP 32700. Email: ionel@inca.edu.cu