Aplicación del sistema de Planta Donante de Micelio (PDM) en la micorrización in vitro de papa
Application of Mycelium Donor Plant (MDP) system on in vitro mycorrhization of potato
Dra.C. Kalyanne Fernández Suárez,I Dr.C. Stéphane Declerck,II Dr.C. Loiret Fernández García,III Dr.C. Eduardo Ortega DelgadoIII
IInstituto Nacional de Ciencias Agrícolas, INCA. Carretera a Tapaste,
km 3½, San José de Las Lajas, Mayabeque. Cuba.
IIEarth and Life Institute – Mycology, Université Catholique de Louvain (UCL), Croix du Sud 2 bte L7.05.06, 1348 Louvain-la-Neuve, Bélgica.
IIIUniversidad de la Habana. Calle 25 esq. J. Plaza de la Revolución.
La Habana. Cuba.
RESUMEN
La micorrización de plantas in vitro es un proceso altamente complejo que depende de garantizar las necesidades de plantas y hongos, organismos que tienen requerimientos nutricionales diferentes. En la actualidad, aún no se ha diseñado un sistema de cultivo in vitro que garantice la micorrización eficiente de plantas de Solanum tuberosum L. (papa), aunque algunos autores han realizado varios intentos por lograrlo. El objetivo de este estudio fue evaluar la aplicabilidad del sistema Planta Donante de Micelio (PDM) a la micorrización in vitro de este cultivo para obtener plantas micorrizadas en un corto periodo, con probada utilidad práctica. Experimentalmente se trabajó con un sistema diseñado de forma similar al PDM en el cual se asociaron plantas de S. tuberosum, de siete días de subcultivadas, con una extensa red de micelio proveniente de plantas de Medicago truncatula Gaertn., previamente micorrizadas con el hongo micorrízico arbuscular Rhizoglomus intraradices. Se obtuvieron elevados niveles de colonización (55 %) de las plantas de papa a solo 12 días de haber entrado en contacto con las redes de micelio. Las plantas de papa se encontraban en condiciones óptimas para ser trasplantadas y fueron capaces de reproducir la colonia fúngica cuando se replantaron en medios frescos, produciendo un elevado número de estructuras extraradicales (micelio y esporas), demostrando la aplicabilidad del sistema PDM en la micorrización in vitro de S. tuberosum.
Palabras clave: cultivo in vitro, redes de micelio extraradical, Rhizoglomus intraradices, Solanum tuberosum.
ABSTRACT
In vitro mycorrhization of plants is a highly complex process that depends on ensuring the needs of plants and fungi, organisms with different nutritional requirements. Today doesn´t exist an in vitro culture system which guarantee the mycorrhization of Solanum tuberosum L. (potato), although some authors have made several attempts to achieve it. The aim of this study was to evaluate the applicability of Mycelium Donor Plant (MDP) system for in vitro mycorrhization of potato plants in order to get mycorrhized plants in a short period, with proven practical use. Experimentally, a similar system to MDP, in which S. tuberosum plants (seven days old) were associated with an extensive network of mycelium coming from Medicago truncatula plants, previously mycorrhized with the AMF Rhizoglomus intraradices was design. High levels of potato plant colonization (55 %), just 12 days after contact the mycelium networks, were obtained. Potato plants were in excellent conditions to be transplanted and they were able to reproduce the fungal colony when they were replanted in fresh media, producing a large number of extraradicals structures (mycelium and spores), demonstrating the applicability of MDP system for in vitro mycorrhization of S. tuberosum.
Key words: extraradical micelium network, in vitro culture, Rhizoglomus intraradices, Solanum tuberosum.
INTRODUCCIÓN
Desde hace aproximadamente tres décadas se han venido desarrollando en Cuba, diferentes líneas de investigación encaminadas al estudio y la introducción de los HMA en los sistemas de producción agrícola, basados en el amplio espectro de la simbiosis micorrízica arbuscular (1, 2). Producto de la importancia de este tema, la búsqueda de nuevas vías de inoculación es objetivo prioritario, siendo la micorrización de plantas in vitro una de las mayores aspiraciones.
La micorrización de plantas in vitro es un proceso altamente complejo, que depende de garantizar las necesidades de plantas y hongos (3), organismos con requerimientos nutricionales diferentes, y en el caso específico de los HMA, sólo se cuenta con escasa información. Los medios de cultivo, los sistemas de cultivo y la fuente de inoculante empleada, son los factores que determinan su éxito (4).
En la actualidad, aún no se ha diseñado un sistema de cultivo in vitro que garantice la micorrización eficiente de plantas de S. tuberosum. Aunque algunos autores (4, 5) han realizado varios intentos por lograrlo y se han utilizado varios sistemas y medios de cultivo, no se ha podido acelerar el proceso de micorrización de las plantas, ni homogenizar la germinación de los propágulos utilizados. Esos autores también han demostrado que el empleo de esporas como inóculo no garantiza la colonización del sistema radical en un periodo corto, ni la producción de un elevado número de estructuras extrarradicales, debido a que estos propágulos requieren de un tiempo prolongado para germinar y colonizar, además se ven influenciados por diversos factores, difíciles de controlar en tales condiciones.
En el año 2009 se publicó un sistema de micorrización in vitro de plantas denominado Planta Donante de Micelio (PDM), el cual explota por primera vez la capacidad colonizativa del micelio extrarradical y en el que se obtienen grandes cantidades de inóculo micorrízico in vitro, a la vez que se logran micorrizar plantas de M. truncatula en condiciones autotróficas (6).
Este sistema se basó en la poca especificidad que muestran las asociaciones micorrízicas en la naturaleza, donde las hifas que emergen de raíces micorrizadas vivas son capaces de conectar plantas de iguales o diferentes especies a través de extensas redes de micelio (6, 7).
Teniendo en cuenta los antecedentes relacionados con la micorrización in vitro de S. tuberosum, se decidió evaluar la aplicabilidad del sistema PDM a la micorrización in vitro de este cultivo para obtener plantas micorrizadas en un corto periodo con probada utilidad práctica.
MATERIALES Y MÉTODOS
En este estudio se utilizó el sistema PDM (6) para la producción rápida y homogénea de plantas de papa micorrizadas in vitro, empleando como fuente de inoculante las redes de micelio extrarradical. Se tuvieron en cuenta resultados previos (4) en los que se demostró, al emplear esporas como inóculo, que éstas no son un propágulo adecuado para micorrizar vitroplantas de papa en tales condiciones, ya que requieren de un tiempo prolongado para germinar y establecer la colonización en las raíces.
Por otra parte, debido a los altos requerimientos nutricionales que tiene la papa y a su rápido desarrollo en condiciones in vitro, es necesario el control de otros factores (ej. nutricionales, ambientales) que hacen complejo el manejo de esos sistemas. Al respecto se hace referencia en un estudio anterior (4) en el que se evaluaron diferentes sistemas y medios de cultivo para garantizar la micorrización in vitro de plantas de papa y se utilizó esporas como inoculante, dejando clara la necesidad de búsqueda de otros propágulos para asegurar la colonización rápida de S. tuberosum.
Material biológico
Se utilizaron vitroplantas de Solanum tuberosum L. cv. Desirée suministradas por los laboratorios de la Estación de Haute, Bélgica. La multiplicación del material vegetal se realizó por micropropagación de los cortes nodales y se colocaron 20 explantes en cajas de cultivo (90 x 60 x 50 mm) estériles, conteniendo 50 mL de medio sólido MS (Murashige y Skoog) (8) (Duchefa, Biochemie) suplementado con 20 g L-1 de sacarosa y 3 g L-1 Gel Gro® (ICN, Biomedicals, Inc., Irvine, USA) como agente gelificante, ajustado a pH 5,7 antes de la esterilización. Las cajas de cultivo fueron transferidas a cámara de crecimiento bajo condiciones controladas (22 °C, fotoperiodo de 16 h día-1 y flujo de fotones fotosintéticos de 225 µmol m-2 s-1), para garantizar el desarrollo de los explantes.
Se trabajó además con semillas de Medicago truncatula Gaertn. cv. Jemalong, cepa A-17 (SARDI, Australia), que se esterilizaron por inmersión en hipoclorito de sodio con 8 % de cloro activo, durante 10 minutos. Se lavaron con agua desionizada estéril y se incubaron en la oscuridad (27 °C) para germinar en placas Petri de 90 mm diámetro, conteniendo 35 mL de medio SRM (Strullu y Romand modificado) (9) sin azúcares y sin vitaminas, solidificado con 3 g L-1 de Gel Gro®. En cada placa se colocaron 25 semillas. Las plantas se utilizaron cuatro días después de la germinación.
El inóculo fúngico utilizado (Rhizoglomus intraradices Schenck & Smith, MUCL-41833), de cuatro meses de cultivo, se adquirió en GINCO (Colección in vitro de Glomeromycota, BCCM/MUCL, Unidad de Microbiología, Universidad católica de Lovaina, Lovaina la Nueva, Bélgica). La cepa fue suministrada en placa Petri de 90 mm diámetro, en asociación con raíces transformadas (Ri T-DNA) de zanahoria (Daucus carotaL.) en medio SRM, solidificado con 3 g L-1 de Gel Gro®. El inóculo se subcultivó utilizando la metodología descrita por Cranenbrouck y colaboradores (10) y se incubó a 27 °C en la oscuridad en posición invertida.
Procedimiento para establecer el sistema PDM
Para establecer el sistema PDM se utilizó como base el sistema publicado por Voets (6), empleando la misma metodología en su diseño. En el compartimento radical de cada placa Petri se colocó igualmente una planta de M. truncatula de cuatro días de germinada. Las pequeñas raíces se colocaron en la superficie del medio de cultivo y el sistema aéreo extendiéndose fuera de la placa.
Se inocularon 50 esporas de R. intraradices sobre las raíces, previa extracción de un cultivo monoxénico de cuatro meses de edad, por solubilización del gel (10), además se continuó con el cierre y sellado de los sistemas (6). Los mismos fueron transferidos a cámara de crecimiento bajo condiciones controladas (22°C, fotoperiodo de 16 h día-1 y flujo de fotones fotosintéticos de 225 µmol m-2 s-1) y mantenidos en posición vertical.
Semanalmente, se añadieron 5 mL de medio SRM a una temperatura aproximada de 35 °C con 4 g L-1 de Gel Gro®. De esta manera se garantizó también un nivel adecuado del medio sobre la pared divisoria (± 2 mm), para facilitar el paso de las hifas hacia el compartimento hifal (CH). Después de ocho semanas de cultivo se determinó el número de hifas cruzando sobre la pared divisoria entre el compartimento radical (CR) y el CH, la longitud del micelio y el número de esporas producidas en el CH.
El número de hifas cruzando sobre la pared divisoria fue cuantificado utilizando un microscopio compuesto de campo brillante (125-250X, Olympus BH-2, Olympus Optical GmbH, Alemania). La longitud micelial y el número de esporas desarrollados en el CH se evaluaron empleando un microscopio de disección (10-40X, Olympus SZ40, Olympus Optical GmbH, Alemania) (5).
Se tomaron dos grupos de 25 unidades experimentales (cada sistema se consideró una unidad experimental) para continuar con la siguiente fase del estudio. Posteriormente, se realizaron dos nuevos orificios (± 2mm diámetro) en el CH de los sistemas, tal como describen otros investigadores (6).
Las placas se abrieron cuidadosamente y se colocaron dos plantas de papa de siete días de subcultivo, presentando aproximadamente 3 cm de longitud y de tres a cuatro hojas por planta. Las raíces se colocaron suavemente sobre el medio de cultivo y el vástago se ubicó hacia el exterior de la placa. Todos los sistemas fueron sellados cuidadosamente, las placas se cubrieron con bolsas de polietileno negro, dejando el sistema aéreo de las plantas expuesto a condiciones de cámara de crecimiento similares a las antes mencionadas, pero en posición horizontal.
Evaluaciones
En este momento del estudio, las 25 unidades experimentales se dividieron en cinco grupos, de manera que cada grupo compuesto por cinco unidades sólo se mantuvo en contacto con la red de micelio los días correspondientes para completar la dinámica. Transcurridos 3, 6, 9, 12 y 15 días, de acuerdo a la dinámica establecida, se extrajo de sus sistemas una planta por unidad experimental, las plantas restantes se utilizaron para el replante. Posteriormente, se tiñeron las raíces de las plantas extraídas (11) y se procedió a estimar la frecuencia e intensidad de la colonización (12).
A los 3, 6, 9, 12 y 15 días de haber entrado en contacto con la red de micelio, las plantas restantes de las cinco réplicas se extrajeron cuidadosamente y se transfirieron a placas Petri de 90 mm diámetro. Las plantas se transfirieron a medio E (MS modificado) (4) sin azúcar ni vitaminas. La producción de esporas por placa Petri y el desarrollo del micelio extrarradical, se midieron semanalmente durante cuatro semanas (5).
Análisis estadístico
Después de comprobada la normalidad y la homogeneidad de varianza (Test de Brown-Forsythe), utilizando el paquete estadístico SPSS Version 21 para Windows (13), los datos fueron sometidos a Análisis de Varianza de clasificación simple. Los valores de las variables frecuencia e intensidad fueron transformados según la expresión arcsen√x/100 y comprobada su distribución normal. Se realizó Análisis de Varianza y Test de Tuckey (14) para formar los dos grupos de 25 sistemas PDM que serían utilizados posteriormente para colonizar las plántulas de S. tuberosum, teniendo en cuenta que no existieran diferencias significativas para p<0,05 en las tres variables evaluadas (número de hifas cruzando la pared divisoria entre compartimentos, longitud micelial en el compartimento hifal y número de esporas en el compartimento hifal). Con los valores de producción de esporas y de longitud del micelio en los distintos tiempos de muestreo (3 a 12 días), en las diferentes semanas después de la asociación, se calcularon los intervalos de confianza de la media al 95 % de probabilidad, atendiendo al número de repeticiones y la reproducibilidad de los datos. Se utilizó igualmente el paquete SPSS Version 21 para Windows (13).
RESULTADOS Y DISCUSIÓN
Entre los 10 y 12 días de comenzado el experimento se observaron los primeros puntos de contacto entre las hifas de germinación provenientes de las esporas y las raíces de las plantas y el micelio comenzó a desarrollarse de manera profusa dentro del medio en los compartimentos radicales. Este comportamiento fue similar al informado en el estudio, en el que se propone el sistema para la micorrización de plantas de M. truncatula, utilizando la misma especie fúngica (6).
Transcurridas seis semanas, las hifas comenzaron a cruzar sobre la pared divisoria entre el CR y el CH, contabilizándose alrededor de 463 ± 28 hifas que habían cruzado al CH al final de este periodo. En ambos compartimentos el micelio se caracterizó por presentar abundantes hifas primarias o corredoras e hifas de bajo orden portando esporas, estructuras ramificadas de absorción (ERA) y esporas asociadas a ERA; comportamiento típico de la colonia de R. intraradices en condiciones in vitro (6, 15).
Dos semanas más tarde, el micelio extrarradical había colonizado intensamente casi la totalidad del CH y se habían producido un promedio de 1 338 ± 186 cm de hifas y más de 1 400 esporas (1 420 ± 238). En las 25 unidades experimentales que se seleccionaron para asociar las plantas de papa, divididas en cinco grupos de cinco sistemas, no se observaron diferencias significativas entre los grupos para las diferentes variables analizadas (Tabla), apreciable en los valores de p.
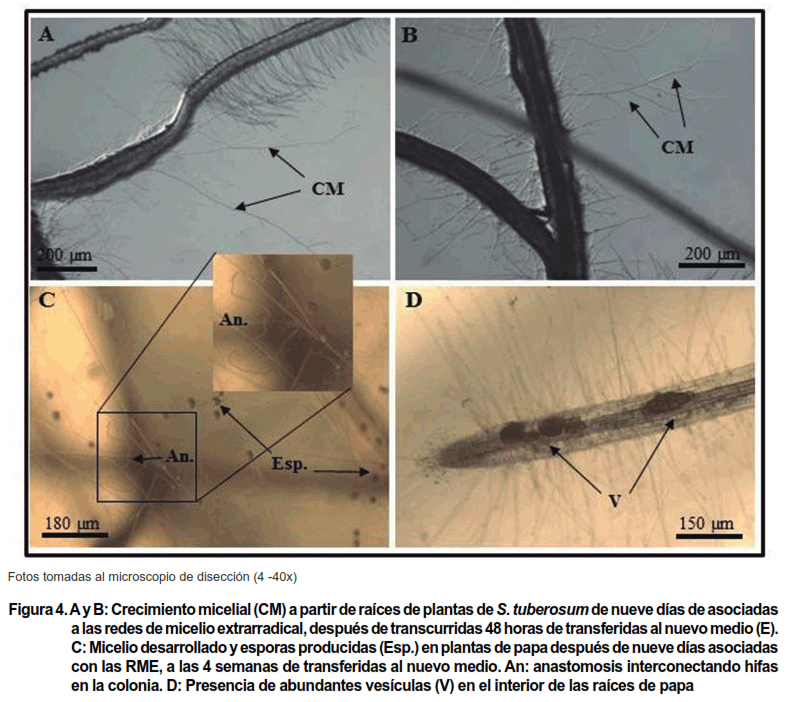
Transcurridos los tres primeros días de haber asociado las plantas de papa a las RME se comenzaron a observar numerosos apresorios en la superficie de las raíces de ambas plantas (Figura 1 A y B), localizados en las zonas apical y subapical. Estas estructuras son las primeras que se observan después del contacto entre la hifa de germinación y la superficie del sistema radical también es la que permite al propágulo infectivo adherirse a la raíz para, a partir de ella, comenzar a penetrar la epidermis y establecerse en el interior radical (16).
Al sexto día se apreciaron algunas vesículas en el interior de las raíces que comenzaron a ser numerosas a partir del noveno día (Figura 1 C y D), siendo más frecuentes en las plantas que estuvieron 12 y 15 días asociadas a los sistemas.
En la Figura 1 E se muestra el estado saludable de las plantas de papa creciendo en medio SRM, después de nueve días de asociadas a la red de micelio presente en el compartimento hifal del sistema PDM. El medio SRM se caracteriza por tener muy bajos contenidos nutricionales (3), ya que está diseñado para garantizar el establecimiento fúngico y no es adecuado para el desarrollo de plantas de papa in vitro; sin embargo, como las plantas estuvieron creciendo solo 12 días en este medio, no se observaron síntomas de deficiencia nutricional, contrario a lo que informaron otros autores (4) cuando inocularon las plantas con esporas en ese mismo medio y el experimento tuvo una duración de 40 días.
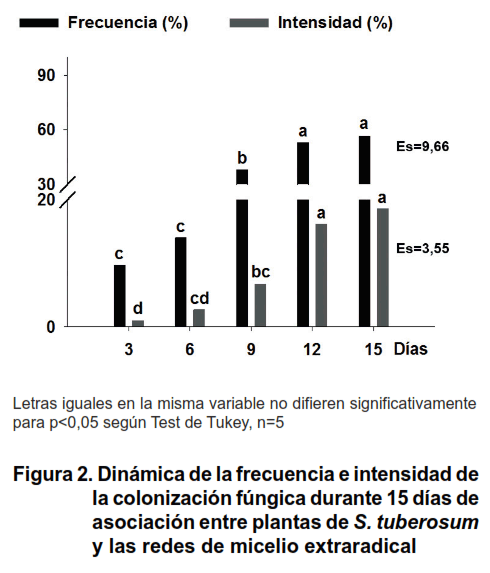
Las evaluaciones realizadas durante los días de asociación de las plantas a las RME (3, 6, 9, 12 y 15 días) que reflejan el establecimiento de la micorrización, se muestran en la Figura 2.
Los niveles de colonización micorrízica, expresados por las variables frecuencia e intensidad, comenzaron a aumentar después de los tres primeros días de asociación con las RME hasta alcanzar un valor máximo, cercano al 60 %, a los 12 días. A partir de ese momento se mantuvieron constantes hasta el final de la dinámica, no mostrando diferencias entre las plantas asociadas a los 12 y 15 días.
Según el análisis realizado en la literatura científica, tan altos valores de colonización no habían sido obtenidos hasta este momento en S. tuberosum en condiciones in vitro (5) y son superiores a los alcanzados por el cultivo en condiciones de campo (17, 18), los cuales no superan el 30 %.
Los altos porcentajes de colonización alcanzados en las plantas se deben a la densa red de micelio compuesto por cientos de hifas en crecimiento activo que emergían de las raíces previamente colonizadas y se dispersaban alrededor de las raíces de las plantas asociadas, tal como describieron los autores del sistema utilizado (6). En el sistema de experimentación, el micelio proveniente de las raíces de las plantas donantes ocupó casi completamente el compartimento hifal, produciendo alrededor de 1340 cm de hifas, por lo que el acceso de estas hifas a las raíces de las plantas recién asociadas resultó extremadamente sencillo.
Aunque los valores que se obtuvieron en este estudio, respecto a la longitud de hifas en el compartimento hifal, son inferiores a los informados por Voets (6), quienes refieren haber cuantificado alrededor de 4 000 cm de hifas, fueron suficientes para que las plantas alcanzaran tan altos valores de colonización en un periodo de tiempo de solo 12 días.
Los resultados difieren de otros publicados previamente (5, 19) en sistemas de cultivo in vitro en los que se inocularon plantas de S. tuberosum y M. truncatula, respectivamente, utilizando una dosis elevada de esporas (100 por placa Petri) como fuente de inóculo. Dichos autores informaron niveles de colonización cercanos a 50 %, transcurridas ocho semanas de cultivo y plantas con severos síntomas de deficiencia nutricional, poniendo de manifiesto lo inconveniente de emplear esporas como inóculo en estos sistemas.
Como se analizó anteriormente, cuando se utilizan esporas como fuente de inóculo el proceso de colonización puede retardarse, no solo en ocurrir, sino también, en extenderse en el sistema radical (4). Esto puede deberse a que las esporas presentan falta de homogeneidad en la germinación, limitada capacidad de crecimiento independiente y los puntos de colonización que originan son locales y están condicionados por la proximidad a la raíz (20, 21).
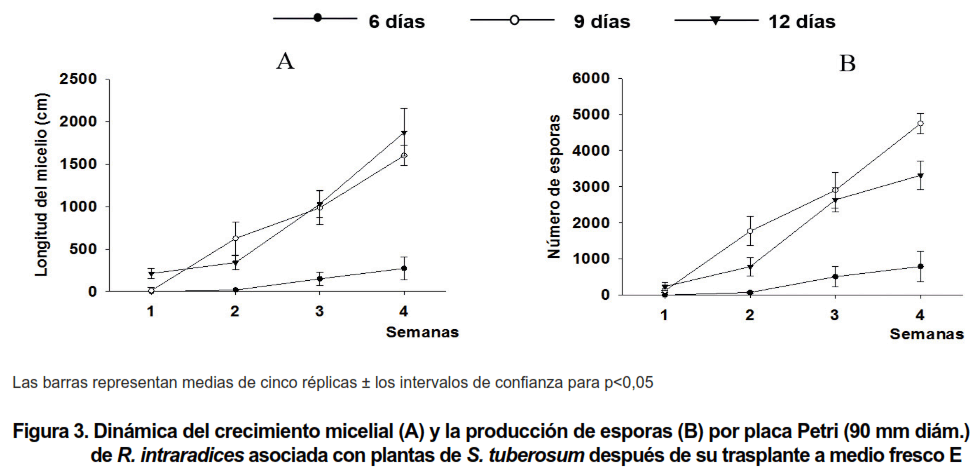
Al extraerse las plantas de papa y transferirse a medio fresco E se pudo observar un comportamiento fúngico diferenciado, en dependencia de los días que habían estado las plantas asociadas con las RME (Figura 3 A y B).
En las plantas que permanecieron solo tres días asociadas a las RME no se observó nuevo crecimiento micelial durante las cuatro semanas de muestreo, mientras que en el caso de las plantas asociadas seis días o más, sí se apreció un marcado crecimiento micelial, proveniente tanto de hifas intactas o dañadas que se encontraban adheridas a la superficie de las raíces, como de hifas que emergían desde el interior radical (6). Este crecimiento fue ganando en intensidad a medida que las plantas estuvieron más tiempo (9 y 12 días) (Figura 3 A y B) en contacto con las redes de micelio y por tanto presentaban un mayor número de estructuras fúngicas intrarradicales.
Se observó que las hifas emergieron desde el interior de las raíces a partir de las 48 horas de trasplantadas, en las plantas que estuvieron 9 y 12 días (Figura 4 A y B) asociadas a las RME y se caracterizaron por un crecimiento recto y profuso dentro del medio.
También se observaron numerosas vesículas en el interior de las raíces (Figura 4 D) y al cabo de la primera semana el micelio se había dispersado intensamente en el medio de cultivo. La producción de esporas estuvo en correspondencia con el desarrollo micelial y alrededor de 100 esporas por placa Petri ya habían sido producidas durante la primera semana de muestreo.
Transcurridas cuatro semanas, el micelio mostraba el crecimiento típico de una colonia de R. intraradices, observándose numerosas anastomosis (Figura 4 C) y un considerable número de esporas por placa Petri (Figura 4 C y Figura 3 B), con valores cercanos a 5 000.
Las plantas que estuvieron 12 días en contacto con las RME mostraron un patrón de crecimiento del nuevo micelio similar al antes descrito para las plantas de nueve días, aunque produjeron un menor número de esporas por placa Petri.
Al analizar los valores de intensidad de la colonización, caracterizada por la presencia de vesículas y arbúsculos se confirmó, que el estado en que se encuentre la micorrización es de vital importancia para garantizar un abundante recrecimiento del micelio. Dichas estructuras solo se observaron en las plantas que estuvieron más de seis días asociadas con la red de micelio y por su parte, los valores de intensidad fueron significativamente mayores a partir de los 12 días de asociación (Figura 2).
En las plantas que se asociaron en el sistema PDM no se pudo relacionar el recrecimiento hifal con la presencia de arbúsculos, debido a que las primeras vesículas también se observaron después de seis días de haber entrado en contacto las raíces de las plantas con las redes de micelio. Estas estructuras de paredes gruesas, además de ser importantes órganos de almacén, juegan un significativo rol como propágulos en fragmentos de raíces (5, 10) y posiblemente también en raíces vivas. Por otra parte, no debe descartarse la posibilidad de que también haya ocurrido recrecimiento micelial a partir de las hifas intrarradicales, ya que las hifas de los HMA tienen la capacidad de crecer hacia el exterior de las raíces y explorar nuevos ambientes (3), como se observó también en este experimento.
En las plantas que se asociaron con las redes de micelio se apreció, además, que las hifas extrarradicales unidas a las raíces (removidas del medio inicial para su transferencia a medios frescos), continuaron creciendo a través de secciones intactas o dañadas. Esto podría representar otro mecanismo de importancia para el nuevo crecimiento fúngico en plantas trasplantadas. Según algunos autores (21), cuando las hifas de los HMA sufren daños de forma natural, son capaces de volver a crecer, repararse, explorar el ambiente circundante y además colonizar nuevas raíces, debido a que cuentan con eficientes mecanismos de reparación.
Otros autores han demostrado también que al cortar y extraer el medio de cultivo de un compartimento hifal y añadir medio fresco, se propicia el crecimiento de nuevo micelio de forma sincronizada, a través del recrecimiento de las hifas cortadas o de la formación de nuevas hifas (22).
El abundante número de esporas (~ 5 000) producido por R. intraradices al colonizar plantas de S. tuberosum (Figura 3 B) también refleja las ventajas del sistema PDM, al informarse en este estudio la mayor cantidad de esporas obtenidas in vitro en un periodo de cuatro semanas en plantas de papa colonizadas, en relación con el resto de los sistemas de cultivo que han sido publicados hasta el momento para esta especie vegetal.
Cuando se utiliza el sistema PDM se pueden obtener plantas de papa micorrizadas a los pocos días de asociadas al sistema (entre 9 y 12 días), con elevados niveles de colonización, capaces de reproducir las colonias fúngicas cuando son trasplantadas a nuevos medios. Además, si se realiza un manejo cuidadoso de los sistemas, evitando posibles contaminaciones microbianas, éstos pueden ser reutilizados permitiendo asociar dos nuevas plantas en cada sistema. Estas plantas micorrizadas a tan corta edad se encuentran en un estadio fisiológico que permite su trasplante a invernadero con el consiguiente beneficio que representa la micorrización en esa fase y la producción masiva de inóculo micorrízico en placas de cultivo o en biorreactores.
CONCLUSIONES
La inoculación de plantas de papa con micelio extrarradical de Rhizoglomus intraradices proveniente de una planta donante, permite obtener elevados niveles de colonización in vitro y reducir el tiempo del proceso colonizativo de S. tuberosum. Estas plantas a solo cuatro semanas del trasplante reproducen la colonia fúngica en medios frescos. Se produce un elevado número de estructuras extrarradicales (micelio y esporas) y el número de esporas producidas es de aproximadamente 100 por centímetro cúbico.
Como ha quedado expuesto a través de este estudio el sistema PDM puede ser adaptado a otras especies de plantas y podría ser aplicado en diversos campos de investigación relacionados con los HMA, no sólo desde el punto de vista de la investigación básica, sino también práctica. Por otra parte, el sistema constituye la base para el desarrollo de un futuro sistema de micorrización de plantas in vitro que permita la introducción de los HMA en las prácticas actuales de producción vegetal y la obtención de plantas in vitro eficientemente micorrizadas, las cuales probablemente estarán más aptas para enfrentar el estrés que representa la adaptación a las condiciones edafoclimáticas.
BIBLIOGRAFIA
1. Golubski, A. J. “Dual plant host effects on two arbuscular mycorrhizal fungi”. Pedobiologia, vol. 54, no. 4, 2011, pp. 209-216, ISSN 0031-4056, DOI 10.1016/j.pedobi.2011.03.003.
2. Shuab, R.; Lone, R.; Naidu, J.; Sharma, V.; Imtiyaz, S. y Koul, K. K. “Benefits of inoculation of arbuscular mycorrhizal fungi on growth and development of onion (Allium cepa) plant”. American-Eurasian Journal of Agriculture & Environmental Sciences, vol. 14, no. 6, 2014, pp. 527–535, ISSN 1990-4053.
3. IJdo, M.; Cranenbrouck, S. y Declerck, S. “Methods for large-scale production of AM fungi: past, present, and future”. Mycorrhiza, vol. 21, no. 1, 2011, pp. 1-16, ISSN 0940-6360, 1432-1890, DOI 10.1007/s00572-010-0337-z.
4. Fernández, S. K.; Fernández, M. F. y Declerck, S. “Búsqueda de un medio de cultivo para la micorrización in vitro de plántulas de papa (Solanum tuberosum L.)”. Cultivos Tropicales, vol. 34, no. 4, 2013, pp. 9-19, ISSN 0258-5936.
5. Voets, L.; Dupré de Boulois, H.; Renard, L.; Strullu, D.-G. y Declerck, S. “Development of an autotrophic culture system for the in vitro mycorrhization of potato plantlets”. FEMS Microbiology Letters, vol. 248, no. 1, 2005, pp. 111-118, ISSN 0378-1097, DOI 10.1016/j.femsle.2005.05.025.
6. Voets, L.; de la Providencia, I. E.; Fernandez, K.; IJdo, M.; Cranenbrouck, S. y Declerck, S. “Extraradical mycelium network of arbuscular mycorrhizal fungi allows fast colonization of seedlings under in vitro conditions”. Mycorrhiza, vol. 19, no. 5, 2009, pp. 347-356, ISSN 0940-6360, 1432-1890, DOI 10.1007/s00572-009-0233-6.
7. Herrera-Peraza, R. A.; Hamel, C.; Fernández, F.; Ferrer, R. L. y Furrazola, E. “Soil–strain compatibility: the key to effective use of arbuscular mycorrhizal inoculants?”. Mycorrhiza, vol. 21, no. 3, 2011, pp. 183-193, ISSN 0940-6360, 1432-1890, DOI 10.1007/s00572-010-0322-6.
8. Murashige, T. y Skoog, F. “A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures”. Physiologia Plantarum, vol. 15, no. 3, 1962, pp. 473-497, ISSN 0031-9317, 1399-3054, DOI 10.1111/j.1399-3054.1962.tb08052.x.
9. Declerck, S.; Strullu, D. G. y Plenchette, C. “Monoxenic Culture of the Intraradical Forms of Glomus sp. Isolated from a Tropical Ecosystem: A Proposed Methodology for Germplasm Collection”. Mycologia, vol. 90, no. 4, 1998, p. 579, ISSN 0027-5514, DOI 10.2307/3761216.
10. Cranenbrouck, S.; Voets, L.; Bivort, C.; Renard, L.; Strullu, D.-G. y Declerck, S. “Methodologies for in Vitro Cultivation of Arbuscular Mycorrhizal Fungi with Root Organs” [en línea]. En: eds. Declerck S., Fortin J. A., y Strullu D.-G., In Vitro Culture of Mycorrhizas, Ed. Springer-Verlag, Berlin/Heidelberg, 2005, pp. 341-375, ISBN 978-3-540-24027-3, [Consultado: 3 de enero de 2017], Disponible en: <http://link.springer.com/10.1007/3-540-27331-X_18>.
11. Rodríguez, J. Y.; Arias, P. L.; Medina, C. A.; Mujica, P. Y.; Medina, G. L. R.; Fernández, S. K. y Mena, E. A. “Alternativa de la técnica de tinción para determinar la colonización micorrízica”. Cultivos Tropicales, vol. 36, no. 2, 2015, pp. 18-21, ISSN 0258-5936.
12. Declerck, S.; Strullu, D. .; Plenchette, C. y Guillemette, T. “Entrapment of in vitro produced spores of Glomus versiforme in alginate beads: in vitro and in vivo inoculum potentials”. Journal of Biotechnology, vol. 48, no. 1-2, 1996, pp. 51-57, ISSN 0168-1656, DOI 10.1016/0168-1656(96)01396-X.
13. IBM Corporation. IBM SPSS Statistics [en línea]. versión 21.0, [Windows], Ed. IBM Corporation, 2012, U.S, Disponible en: <http://www.ibm.com>.
14. Tukey, J. W. “Bias and confidence in not quite large samples”. The Annals of Mathematical Statistics, vol. 29, no. 2, junio de 1958, pp. 614-623, ISSN 0003-4851, DOI 10.1214/aoms/1177706647.
15. Calvet, C.; Camprubi, A.; Pérez-Hernández, A. y Lovato, P. E. “Plant growth stimulation and root colonization potential of in vivo versus in vitro arbuscular mycorrhizal inocula”. HortScience, vol. 48, no. 7, 2013, pp. 897–901, ISSN 1805-9333.
16. Willis, A.; Rodrigues, B. F. y Harris, P. J. C. “The Ecology of Arbuscular Mycorrhizal Fungi”. Critical Reviews in Plant Sciences, vol. 32, no. 1, 2013, pp. 1-20, ISSN 0735-2689, 1549-7836, DOI 10.1080/07352689.2012.683375.
17. Fasi, W.; Wanfu, W.; Yantian, M.; Yongjun, L.; Xiaojun, M.; Lizhe, A. y Huyuan, F. “Prospect of beneficial microorganisms applied in potato cultivation for sustainable agriculture”. African Journal of Microbiology Research, vol. 7, no. 20, 2013, pp. 2150-2158, ISSN 1996-0808, DOI 10.5897/AJMR12x.005.
18. Lone, R.; Shuab, R.; Sharma, V.; Kumar, V.; Mir, R. y Koul, K. K. “Effect of Arbuscular Mycorrhizal Fungi on Growth and Development of Potato (Solanum tuberosum) Plant”. Asian Journal of Crop Science, vol. 7, no. 3, 2015, pp. 233-243, ISSN 1994-7879, DOI 10.3923/ajcs.2015.233.243.
19. Dupré, de B. H.; Voets, L.; Delvaux, B.; Jakobsen, I. y Declerck, S. “Transport of radiocaesium by arbuscular mycorrhizal fungi to Medicago truncatula under in vitro conditions”. Environmental Microbiology, vol. 8, no. 11, 2006, pp. 1926-1934, ISSN 1462-2920, DOI 10.1111/j.1462-2920.2006.01070.x.
20. Smith, S. E. y Smith, F. A. “Fresh perspectives on the roles of arbuscular mycorrhizal fungi in plant nutrition and growth”. Mycologia, vol. 104, no. 1, 2012, pp. 1-13, ISSN 0027-5514, 1557-2536, DOI 10.3852/11-229.
21. Gulbis, N.; Boyer, L. R. y Robinson, G. “Studying the microbiome of AMF cultivated in vitro”. Aspects of Applied Biology, vol. 120, 2013, pp. 71–76, ISSN 0265-1491.
22. Cano, C.; Dickson, S.; González-Guerrero, M. y Bago, A. “In vitro Cultures Open New Prospects for Basic Research in Arbuscular Mycorrhizas” [en línea]. En: ed. Varma A., Mycorrhiza, Ed. Springer, Berlin, Heidelberg, 2008, pp. 627-654, ISBN 978-3-540-78824-9, [Consultado: 3 de enero de 2017], Disponible en: <http://link.springer.com/10.1007/978-3-540-78826-3_30>.
Recibido: 14/12/2015
Aceptado: 07/09/2016
Dra.C. Kalyanne Fernández Suárez, Instituto Nacional de Ciencias Agrícolas, INCA. Carretera a Tapaste, km 3½, San José de Las Lajas, Mayabeque. Cuba. Email: kalyanne@inca.edu.cu