Aspersión foliar con biobras-16 estimula el crecimiento de plantas jóvenes de arroz (Oryza sativa L.) sometidas a tratamiento con NaCl
Biobras-16 foliar spraying enhances rice (Oryza sativa L.) young plant growth under NaCl treatment
M.Cs. Yanelis Reyes Guerrero, Lisbel Martínez González, Dra.C. Miriam Núñez Vázquez
Instituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, provincia Mayabeque, CP 32700.
Resumen
La salinidad es una de las causas de estrés más acuciantes en la actualidad y los brasinoesteroides son compuestos que, además, de estimular el crecimiento vegetal aumentan la tolerancia de las plantas al estrés salino. En el presente trabajo, se evaluó el efecto de la aspersión foliar con un análogo de brasinoesteroides (Biobras-16, BB-16), en plántulas de arroz sometidas a este tipo de estrés. Para ello, se asperjaron plantas jóvenes de arroz del cultivar J-104 con BB-16 (0; 0,01; 0,1; 1 µmol L-1), la mitad de ellas se trataron con NaCl (100 mmol L-1) por siete días, y posteriormente, se transfirieron a solución nutritiva para evaluar su recuperación durante catorce días. Las evaluaciones de crecimiento (longitudes de la parte aérea y de las raíces y masas secas de la parte aérea y de las raíces) se realizaron al inicio y final del tratamiento salino y a los siete y catorce días de recuperación. Partiendo de los resultados de este experimento, se seleccionó la mejor concentración de BB-16 y se ejecutó un segundo experimento, siguiendo la misma metodología, donde, además, de evaluar los indicadores de crecimiento, se determinaron algunos indicadores bioquímicos como malondialdehído, prolina, clorofilas y las actividades de las enzimas peroxidasa y catalasa. Los resultados demostraron que, de forma general, la aspersión foliar con BB-16 a 0,1µmol L-1 estimuló la longitud de las plántulas de arroz en condiciones salinas lo que parece estar asociado a un incremento de las defensas antioxidantes, así como a un aumento de la concentración de clorofilas, fundamentalmente clorofila a.
Palabras clave: brasinoesteroides, catalasas, clorofilas, estrés salino, prolina.
ABSTRACT
Salinity is one of the most pressing stresses today and brassinosteroids are compounds which, not only stimulate plant growth but also improve plant tolerance to salt stress. In this paper, the effect of foliar spraying with a brassinosteroid analogue (Biobras-16, BB-16) on rice seedlings under this kind of stress was evaluated. For this purpose, rice young plants cv. J-104 were sprayed with BB-16 (0; 0,01; 0,1; 1 µmol L-1), half of them was treated with NaCl (100 mmol L-1) for seven days and later, these plants were transferred to nutrient solution to assess recovery during fourteen days. The growth evaluations (shoot and root length and dry mass) were made at the beginning and the end of saline treatment and seven and fourteen days after the recovery. From the results of this experiment, the best BB-16 concentration was chosen and a second experiment, following the same methodology, was performed and, where, the growth indicators and some biochemical indicators such as malondialdehyde, proline, chlorophylls and the activities of peroxidase and catalase enzymes were determined. Results demonstrated, in general, that the foliar spraying with BB-16 0,1 µmol L-1 increased the length of rice seedlings under saline conditions what appears to be associated with increased antioxidant defenses as well as an increase of the concentration of chlorophylls, chlorophyll a fundamentally.
Key words: brassinosteroids, catalases, chlorophylls, salt stress, proline.
INTRODUCCIÓN
La salinidad es una de las causas de estrés más acuciantes en la actualidad. La inadecuada irrigación de los suelos, así como el cambio climático hacen que este fenómeno alcance un nivel global (1). En nuestro país, la superficie agrícola está afectada en un 14 % y otro 15 % más presenta peligros potenciales de salinizaciónA.
Para enfrentar esta problemática, se trabaja en la búsqueda de soluciones que permitan incrementar la productividad de los cultivos en estas condiciones, para lo cual no sólo se pudieran utilizar cultivares tolerantes, sino también trabajar en la introducción de tecnologías que permitan minimizar los daños que estos tipos de estrés provocan en las plantas.
Una de las tecnologías para incrementar los rendimientos agrícolas que se desarrolla en los últimos años, se basa en el uso de productos naturales inocuos desde el punto de vista ecológico, como los brasinoesteroides (BRs).
Estos compuestos son potentes reguladores del crecimiento vegetal de naturaleza esteroidal. Estas hormonas tienen efectos pleiotrópicos como son: estimulación del alargamiento celular y desdiferenciación de protoplastos, regeneración de la pared celular, regulación de la diferenciación de elementos traquearios e incremento de la biomasa y del rendimiento (2). En los últimos años, varios autores han estudiado la protección que los BRs le confieren a las plantas ante determinados tipos de estrés tanto bióticos como abióticos (3, 4).
En Cuba, se investiga en la síntesis y actividad biológica de algunas formulaciones, que tienen como ingredientes activos análogos espirostánicos de brasinoesteroides (5).
Desde el punto de vista práctico, la importancia del uso de análogos cubanos en lugar de compuestos naturales es que estos últimos son más costosos y sus efectos son de corta duración en las plantas (6). En cambio, los análogos, sintetizados en Cuba, han demostrado ser productos más baratos e inocuos con utilidad práctica para incrementar los rendimientos agrícolas entre un 10 y 25 % en diferentes especies, incluyendo Oryza sativa, especie de gran importancia económica para nuestro país; por ser un alimento básico para la población (7). Sin embargo, hasta donde se conoce, no se ha logrado la optimización de la aplicación de estos compuestos en la protección de los cultivos sometidos a diferentes estreses ambientales, debido fundamentalmente, a que se desconoce el mecanismo en que ellos actúan en las plantas en estas condiciones. El conocimiento del modo de acción de los brasinoesteroides en el cultivo del arroz ante condiciones de estrés salino permitiría, en un futuro, una aplicación más efectiva y racional de estos reguladores del crecimiento. Por esta razón, el objetivo de este trabajo es determinar si la aspersión foliar con Biobras-16 es capaz de proteger a plantas jóvenes de arroz de los efectos adversos que provoca el estrés salino, así como evaluar algunos indicadores bioquímicos asociados a esta respuesta.
MATERIALES Y MÉTODOS
Para dar cumplimiento al objetivo propuesto, se utilizaron semillas de arroz (Oryza sativa L.) del cultivar Jucarito-104 (J-104) considerado como susceptible al estrés salinoB. Las semillas se desinfectaron con hipoclorito de sodio comercial durante cinco minutos; posteriormente, se lavaron seis veces con agua destilada estéril y luego se sumergieron en agua durante 24 horas.
Transcurrido este tiempo, se colocaron en placas de Petri, a temperatura ambiente en la oscuridad, para propiciar la germinación. Luego de 48 horas, las semillas germinadas se trasplantaron a vasos plásticos de 240 mL que contenían suelo Hidromórfico Gley Nodular Petroférrico (8) proveniente de la UCTB de Los Palacios, Pinar del Río. Las características de este suelo se encontraban en el rango que permitían el crecimiento adecuado de las plantas de arroz (Tabla I). Las determinaciones se realizaron en el Laboratorio de Análisis Químico del Instituto Nacional de Ciencias Agrícolas (9).
Se utilizaron 20 vasos (cada uno con tres plantas en crecimiento) y se colocaron en cestas plásticas que contenían agua corriente. Las cestas se ubicaron dentro de un umbráculo para evitar el efecto de las precipitaciones.
En el momento de emergencia de la tercera hoja verdadera, se realizó la aspersión foliar con Biobras-16 a las concentraciones de 0,01; 0,1 y 1 μmol L-1. Se asperjó un volumen total de 60 mL para 120 plantas. A todas las soluciones se le adicionó Tween 20 al 0,01 % como detergente. En ese momento, las plantas se dividieron en dos grupos, uno se colocó en cestas a las cuales se les adicionó solución nutritiva Hoagland diluida 1:1 v/v (SN) y el otro se colocó en cestas que contenían solución nutritiva Hoagland diluida suplementada con NaCl 100 mmol L-1 (SN+NaCl).
A los siete días, las plantas que se sometieron a tratamiento salino, se colocaron en solución nutritiva Hoagland diluida, durante 14 días, para su recuperación.
Las evaluaciones de crecimiento (longitud de la parte aérea, longitud de las raíces, masa seca de la parte aérea y de las raíces) se realizaron a 12 plantas por tratamiento al iniciar y finalizar el tratamiento salino y a los siete y 14 días de recuperación.
El experimento se realizó en dos épocas del año, el primero se inició el 24 de diciembre del 2009 y el segundo el 25 de mayo del 2010. En ambos casos se registraron las temperaturas mínimas, medias y máximas y las humedades relativas medias del aire durante las siguientes etapas: crecimiento inicial de las plántulas (desde el trasplante hasta la aplicación del tratamiento con NaCl), período de tratamiento con NaCl (siete días) y fase de recuperación (14 días) (Tabla II). Estos datos fueron tomados de la Estación Meteorológica de Tapaste ubicada aproximadamente a 500 m del área donde se desarrollaron los experimentos.
A partir de los resultados del experimento anterior, se ejecutó un segundo experimento, en los meses de abril-mayo de 2011, donde se utilizó la misma metodología que en el experimento anterior, con la diferencia que la aspersión foliar se realizó con Biobras-16 0,1 μmol L-1. Las evaluaciones de crecimiento (longitudes de la parte aérea y de las raíces, masas secas de la parte aérea y de las raíces) se realizaron a 12 plantas por tratamiento al iniciar y finalizar el tratamiento salino y a los 14 días de recuperación. En el caso de las evaluaciones bioquímicas, realizadas a las hojas de las plantas, éstas se efectuaron al finalizar el tratamiento salino y a los 14 días de recuperación. Se determinaron los siguientes indicadores: prolina según el método de la ninhidrina (10), malondialdehído (MDA) por el método del ácido tiobarbitúrico (11) y clorofilas a, b y totales por extracción con acetona al 80 % (12); así como las actividades de las enzimas antioxidantes peroxidasa (EC 1.11.1.7) (13) y catalasa (EC 1.11.1.6) (14). Además se determinaron las proteínas totales según el método de Bradford (15).
En todos los experimentos se utilizó un diseño completamente aleatorizado. Luego de comprobar que los datos obtenidos cumplieron con los supuestos teóricos de normalidad y homogeneidad de varianza, se les realizó un ANOVA de clasificación simple. En los casos en que hubo diferencias significativas entre tratamientos se realizó una comparación de medias mediante la prueba de Tukey con p ≤ 0,05 (16), utilizando el programa estadístico STATGRAPHICS Plus 5.1 (17).
RESULTADOS Y DISCUSIÓN
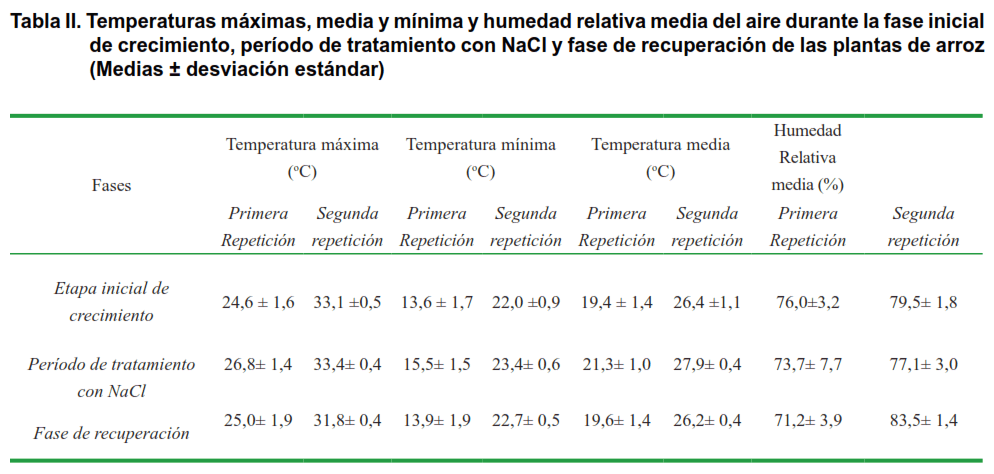
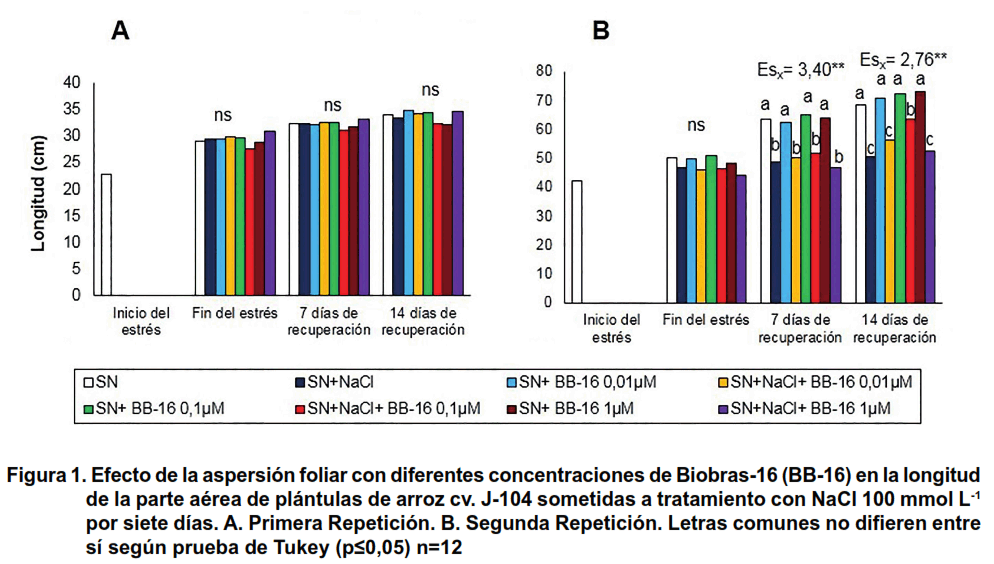
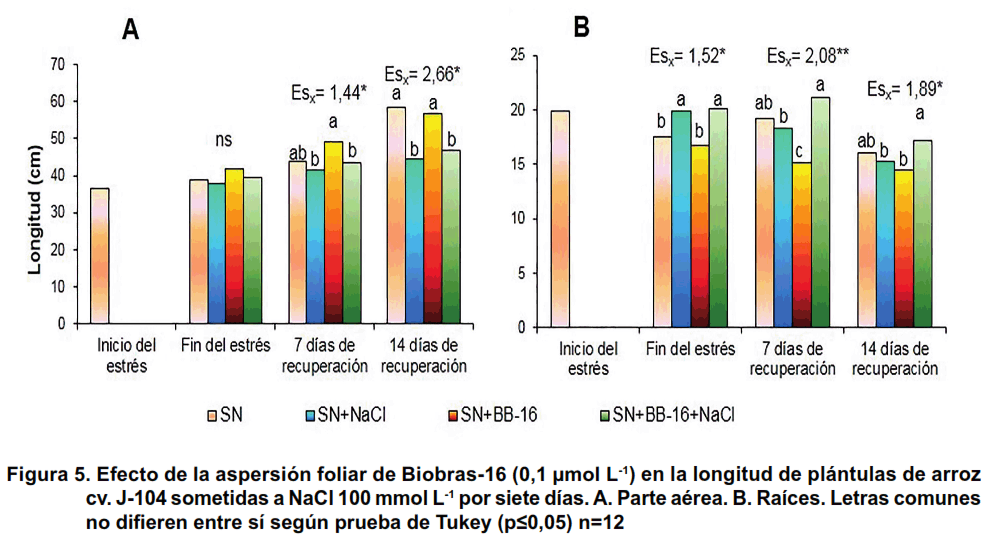
En la Figura 1 se muestra el comportamiento de la longitud de la parte aérea de las plántulas. Como se puede observar, en la primera repetición del experimento, el tratamiento con NaCl no afectó este indicador en ninguno de los momentos evaluados, ni se observó influencia de la aspersión foliar con BB-16. Sin embargo, en la segunda repetición, efectuada en los meses de mayo-junio, se observó que el tratamiento con NaCl produjo una disminución significativa de la longitud de la parte aérea en todos los tratamientos, que se manifestó durante la recuperación. La aspersión foliar con BB-16 0,1 μmol L-1 fue capaz de proteger a las plántulas, estimulando de forma significativa la longitud de la parte aérea a los 14 días de la recuperación.
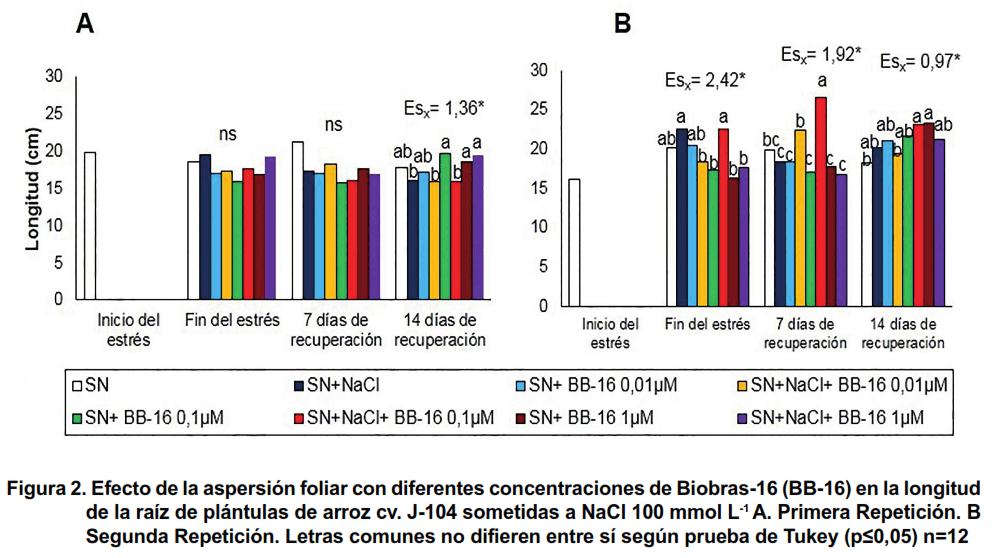
En la longitud de las raíces se observa que, en la primera repetición, de forma general, el tratamiento con NaCl no afectó este indicador en ninguno de los tratamientos ni momentos evaluados (Figura 2A), mientras que en la segunda (Figura 2B) se observó que la salinidad provocó un incremento significativo de este indicador en las plántulas asperjadas con 0,1 μmol L-1 de BB-16, a los siete días de recuperación.
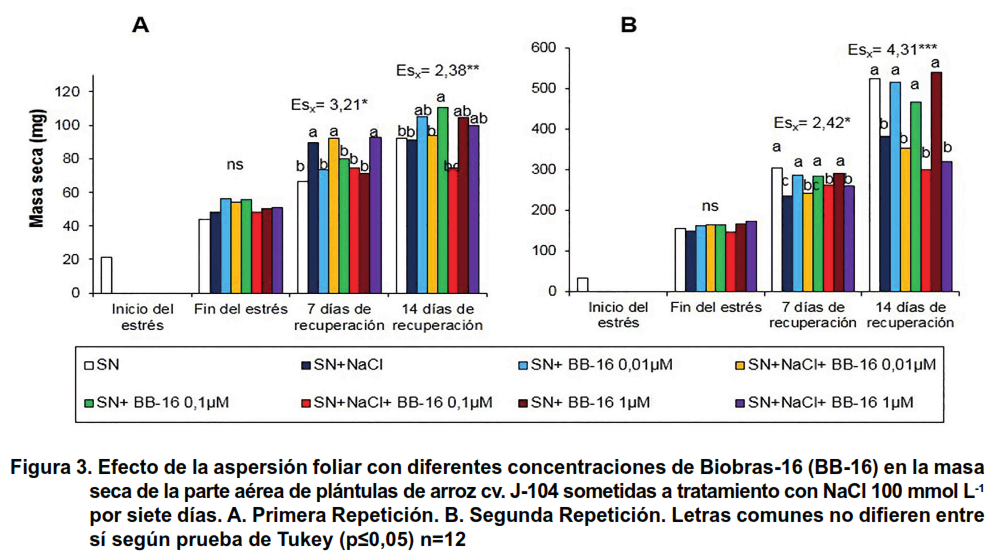
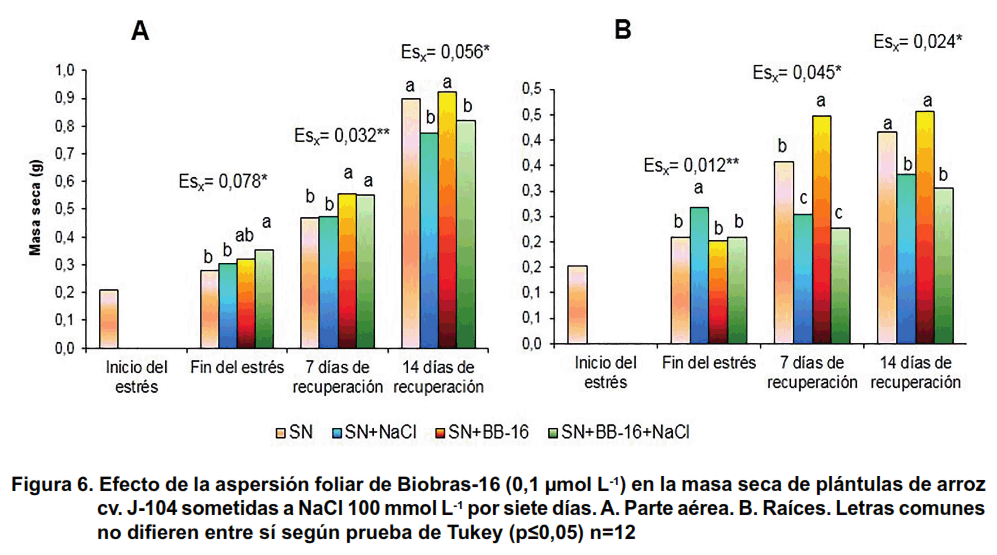
En cuanto a la masa seca de la parte aérea, no se encontraron afectaciones en este indicador una vez finalizado el tratamiento con NaCl en ninguna de las dos repeticiones del experimento. Sin embargo, a los siete días de recuperación en las plántulas con el tratamiento NaCl aumentó de forma significativa la masa seca de las mismas en la primera repetición, y solamente las concentraciones de 0,01 y 1 μmol L-1 de BB-16 conservaron este efecto. No obstante, en la segunda repetición se observó una reducción significativa de este indicador durante la recuperación, excepto en los tratamientos de 0,1 y 1 μmol.L-1, a los siete días de recuperación (Figura 3).
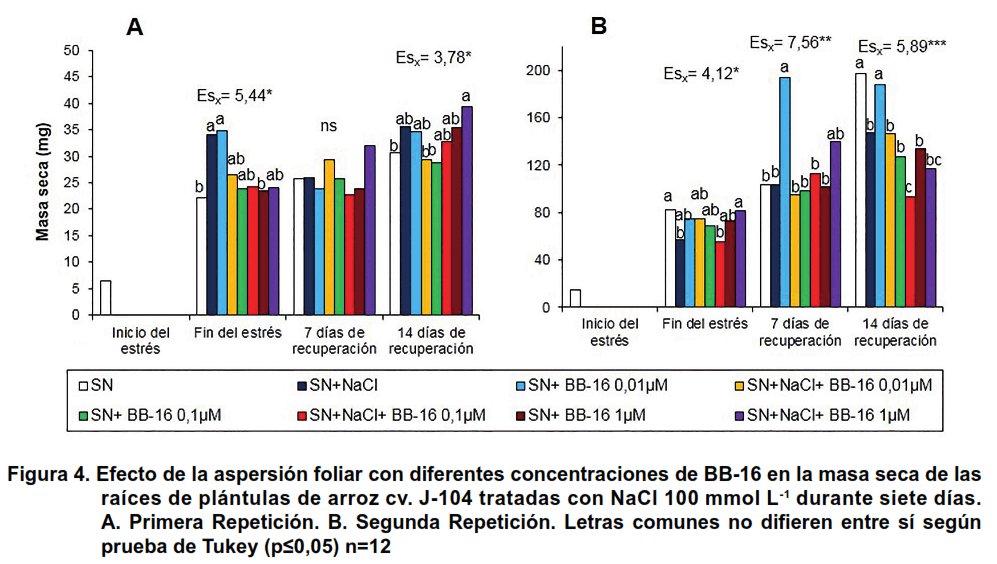
Al evaluar la masa seca de las raíces, en el estrés salino se obtuvo un aumento significativo de este indicador en la primera repetición (Figura 4A), que no fue observada en las plantas asperjadas con BB-16. El comportamiento en la segunda repetición fue diferente, ya que la salinidad disminuyó significativamente la masa seca de las raíces en las plantas controles al finalizar el período de tratamiento con NaCl y a los 14 días de la recuperación; sin embargo, las plantas asperjadas con BB-16 1 μmol L-1 no modificaron significativamente los niveles de masa seca, una vez finalizado el estrés ni durante la recuperación (Figura 4B).
La salinidad afectó negativamente la longitud de la parte aérea, pero esta afectación se produjo en la recuperación, donde se observó una marcada reducción del crecimiento de las plántulas. Es bien conocido que la salinidad inhibe el crecimiento de las plantas y en muchos casos la parte aérea es más afectada que las raíces. Este efecto del estrés salino en el crecimiento es ampliamente conocido ya que tanto el estrés hídrico como la acumulación de iones retrasan los procesos de división y diferenciación celular (18).
Esta disminución en el crecimiento de las plantas tratadas, durante la recuperación, pudiera deberse a que los efectos tóxicos de la acumulación de sales tardan más en manifestarse pero permanecen en el tiempo, aún después de pasar al período de recuperación. Para corroborar esta hipótesis, se ha señalado el hecho de que una vez concluido el período de estrés y sometidas las plantas durante siete días al tratamiento control, estas no lograron recuperarse, lo que demuestra el carácter irreversible de los daños ocasionados (19).
Se apreció que la aspersión foliar con BB-16 0,1 mg L-1 estimuló la longitud de la parte aérea, 14 días después de la recuperación, en comparación al resto de los tratamientos estudiados.
Es de destacar, la diferencia observada en la respuesta de las plantas ante el tratamiento con NaCl en las dos repeticiones efectuadas, la cual pudiera estar relacionada con las condiciones meteorológicas imperantes durante la ejecución del experimento. Como se aprecia en la Tabla II, en el experimento iniciado en diciembre (primera repetición), tanto la temperatura máxima, como la mínima y la media, durante la ejecución del experimento, fue aproximadamente de 7 a 8 ºC más baja que en el iniciado en mayo (segunda repetición) y es bien conocido, el efecto que la temperatura ejerce en el crecimiento del arroz, pues la temperatura óptima para la fase vegetativa de este cultivo es de 25-30 ºC y temperaturas menores ocasionan un retraso en el crecimientoC.
Lo anterior demuestra que las afectaciones de la salinidad en las plántulas también dependen de otros factores, puesto que en el experimento iniciado en el mes de diciembre, donde la temperatura fue más baja, las plantas controles mostraron un menor crecimiento enmascarando el efecto provocado por el estrés salino. Además las bajas temperaturas también pueden haber disminuido la absorción de agua, y por ende de NaCl, en las plántulas de arroz.
En las Figuras 5 y 6 se presentan los resultados de la influencia de la aspersión foliar con Biobras-16 (0,1 µmol L-1) en los indicadores de crecimiento de las plántulas de arroz sometidas o no a tratamiento con NaCl 100 mmol L-1 durante siete días. Se pudo observar que al finalizar el tratamiento con NaCl, no se afectó ninguno de los indicadores del crecimiento evaluados; sin embargo, las plantas que se colocaron en medio salino exhibieron, de forma general, un menor crecimiento después de catorce días de recuperación. Así, en la Figura 5 se muestran las longitudes de la parte aérea y de las raíces. Se aprecia un efecto protector del BB-16 en la longitud de las raíces (Figura 5B), confirmando lo encontrado en la segunda repetición del experimento anterior (Figura 2B).
En cuanto a la masa seca (Figura 6), hubo un efecto protector del BB-16 en la parte aérea, donde la aspersión del compuesto incrementó la masa seca durante el período de estrés y a los siete días de recuperación, en comparación con las plantas que crecieron en solución nutritiva (Figura 6A). Es interesante señalar, que la salinidad incrementó la masa seca de las raíces al finalizar el período de estrés, mientras que la aspersión foliar con BB-16 no modificó este indicador en las plantas tratadas o no con NaCl; sin embargo, durante la recuperación se apreció una reducción significativa de la masa seca de las raíces de las plantas sometidas a tratamiento con NaCl independientemente de que fueran asperjadas o no con BB-16 (Figura 6B).
Este último resultado, que muestra el aumento de la biomasa de la raíz al finalizar el tratamiento salino, se repite en varios experimentos y se han encontrado varios autores que observan un comportamiento similar (20). A pesar de que las raíces se encuentran en contacto directo con la sal, su crecimiento se ve menos afectado que el de la parte aérea, aumentando la razón raíz: parte aérea (18). Esto pudiera ser una adaptación de la planta para aumentar el área de absorción de agua y disminuir el área de transpiración. Sin embargo, en el período de recuperación, la biomasa de la raíz disminuye en las plántulas que fueron previamente estresadas sugiriendo que los efectos a largo plazo de la salinidad afectan la velocidad de crecimiento de este órgano.
Por otra parte, se debe señalar que se ha informado la efectividad del tratamiento a las semillas con BB-16 0,005 mg L-1 para revertir los efectos adversos que la salinidad provocó en el crecimiento inicial de las plantas de los cultivares de arroz J-104 e INCA LP-7, mientras que la respuesta de otro análogo (BB-6) dependió del cultivar (7).
Más recientemente, se evaluó la influencia del tratamiento a las semillas con varias concentraciones de BB-16 y 24-epibrasinólida (EBL), un brasinoesteroide natural, en el crecimiento de las plántulas de dos cultivares de arroz (J-104 y Ginés) en medio salino. Tanto la EBL como el BB-16 fueron capaces de revertir parcialmente la inhibición que en el crecimiento de las plántulas de ambos genotipos de arroz provocó la presencia de NaCl; siendo más efectivos los tratamientos en el cultivar tolerante Ginés (21).
Como se puede apreciar, los resultados mostrados en la literatura, se refieren al tratamiento a la semilla como modo de aplicación, por lo que estos constituyen los primeros resultados que muestran que la aspersión foliar del Biobras-16 pudiera proteger a las plantas jóvenes de arroz del estrés salino.
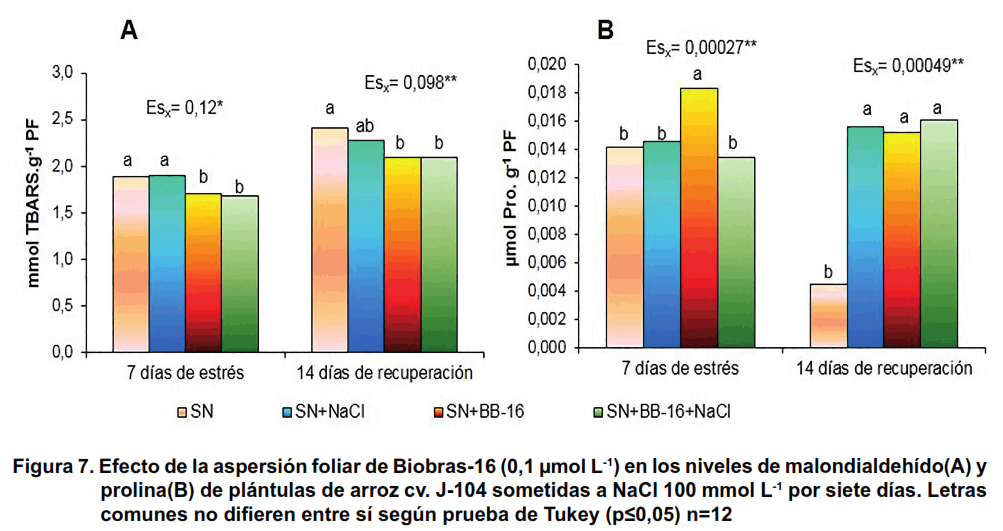
El estrés salino no modificó la concentración de malondialdehído (MDA) en las plántulas en ninguno de los dos momentos evaluados; no obstante, la aspersión foliar con BB-16 disminuyó la concentración de este compuesto con respecto a las plantas controles (Figura 7A). El MDA es uno de los productos finales de bajo peso molecular de la descomposición de los hidroperóxidos lipídicos. Este compuesto es el más utilizado como medida de la peroxidación lipídica en la membrana celular, como consecuencia directa de la acción de las especies reactivas de oxígeno.
Los resultados de este trabajo indicaron que el tratamiento con NaCl durante siete días no provocó daño oxidativo en las membranas de las células de las plantas de este cultivar de arroz hasta los 14 días de recuperación y la aspersión con BB-16 no modificó este comportamiento.
En cuanto a la prolina, el tratamiento con NaCl no modificó la concentración de este metabolito en las plantas controles al final del mismo; sin embargo, la aspersión de BB-16 aumentó los niveles de este aminoácido en las plántulas no estresadas. Por otra parte, en la recuperación se evidenció el incremento en la concentración de este metabolito producto del tratamiento salino y las plantas asperjadas con BB-16 presentaron valores similares de prolina en sus hojas, independientemente, que hubieran sido tratadas o no con sal (Figura 7B).
Otros autores, han observado que la EBL logró disminuir el eflujo de electrolitos y la peroxidación lipídica en plántulas de frijol y pepino expuestas a estrés salino, lo que favoreció el índice de estabilidad de las membranas (22, 23). Algo similar se ha informado en la literatura en el caso de plantas de trigo sometidas a estrés salino y asperjadas con diferentes concentraciones de EBL (24). Igualmente, en plántulas de frijol sometidas a estrés salino la aspersión foliar con EBL aumentó la concentración de prolina tanto en las hojas como en las raíces (22). Sin embargo, la concentración de este aminoácido disminuyó en hojas de plantas de arroz estresadas, cuyas semillas fueron tratadas con EBL (25) y en semillas de frijol tratadas con EBL (5 µmol L-1) y sometidas a estrés salino por ocho días, con respecto a las plántulas estresadas (26).
Aunque la acumulación de prolina se ha observado en arroz expuesto a estrés salino, hoy está claro que este aminoácido tiene múltiples funciones. Numerosos autores plantean que los cultivares más susceptibles a la salinidad tienen mayor contenido de prolina que los cultivares tolerantes (27, 28). Por esta razón, la acumulación de prolina se correlaciona con el grado de deterioro inducido por el estrés (29) y, hay autores que plantean que el aumento de este aminoácido podría deberse auna necesidad de la planta de eliminar el exceso de amonio (30).
Por otro lado, recientemente, se ha descrito el posible papel de la prolina como un antioxidante no enzimático; ya que es capaz de comportarse como aceptor final de los radicales libres como el oxígeno singlete y el radical hidroxilo, por lo que la acumulación de prolina en el tejido puede ser importante en prevenir el daño oxidativo causado por las especies reactivas de oxígeno (31). La prolina, además, estabiliza las membranas y los componentes subcelulares, incluyendo el complejo II de la cadena de transporte electrónico mitocondrial, y estabilizador del potencial redox por reaprovisionamiento de NADP+ (32). Se ha demostrado que la aplicación exógena de prolina proporciona una acción protectora contra el daño oxidativo inducido por el estrés salino. Existen informes que plantean que la aplicación de prolina fue capaz de disminuir la concentración de H2O2 y aumentar la actividad de la enzima ascorbato peroxidasa (APX) en dos cultivares de arroz que difieren en su tolerancia a la salinidad, mientras que también aumentó la actividad de las enzimas peroxidasa y catalasa en el cultivar sensible, tanto en el periodo de estrés como en la recuperación (33). Otros autores, también han demostrado que la aplicación de prolina exógena disminuye el daño oxidativo e incrementa las defensas antioxidantes en diferentes cultivos bajo estrés salino (34, 35).
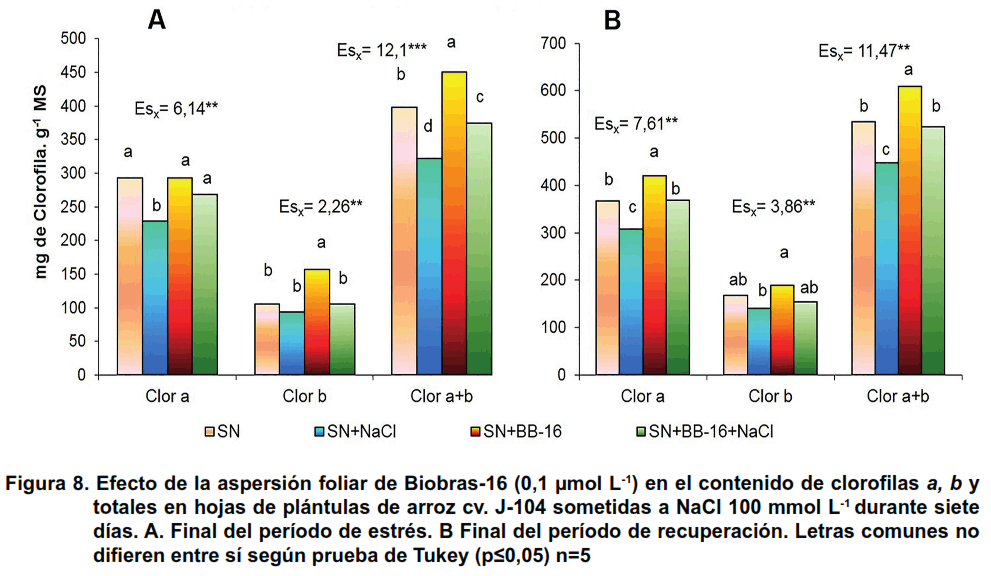
Al analizar los resultados sobre los pigmentos clorofílicos, al final del tratamiento con NaCl, se observó una disminución de la clorofila a y de las clorofilas totales en las plántulas estresadas; mientras que la aspersión foliar con esta concentración de BB-16 fue capaz de contrarrestar este efecto (Figura 8A). En la recuperación, este comportamiento se mantuvo en el caso de la clorofila a y por ende, en las clorofilas totales (Figura 8B). Este resultado concuerda con diferentes autores que han observado la disminución de la concentración de este pigmento ante estrés salino en numerosos cultivos como: lechuga, arroz, pepino y trigo (23, 27, 28, 36).
La reducción en el contenido de clorofila en las hojas en condiciones de estrés salino ha sido atribuida a varias causas: la destrucción de los pigmentos clorofílicos por el incremento de la actividad clorofilasa y la supresión de enzimas de la biosíntesis como la porfobilinógeno desaminasa. También puede estar ocasionada por la interferencia de los iones salinos con la síntesis de novo de proteínas y componentes estructurales de la clorofila (37).
La estimulación de la concentración de clorofila a en las hojas de las plantas asperjadas con BB-16 y sometidas al tratamiento con NaCl, confirma lo obtenido por otros autores quienes han informado que el tratamiento a las semillas con brasinólida, 24-epibrasinólida y homobrasinólida (3 µmol L-1) disminuyó la pérdida de los pigmentos fotosintéticos provocada por el estrés salino (NaCl 150 mmol L-1) en plántulas de arroz (3).
En otros cultivos, también se demostró el efecto de los BRs en la restitución de las moléculas de clorofila ante estrés salino. Por ejemplo, en Phaseolus vulgaris y Hordeum vulgare el pretratamiento a las semillas con brasinoesteroide (5 µmol L-1) revirtió la disminución del contenido de clorofila provocado por el estrés salino (150 mmol L-1) alcanzando niveles similares a las plantas controles (26). De igual forma, la aspersión foliar de EBL en plantas de frijol sometidas a estrés salino revirtió la disminución en las clorofilas provocadas por este estrés (22).
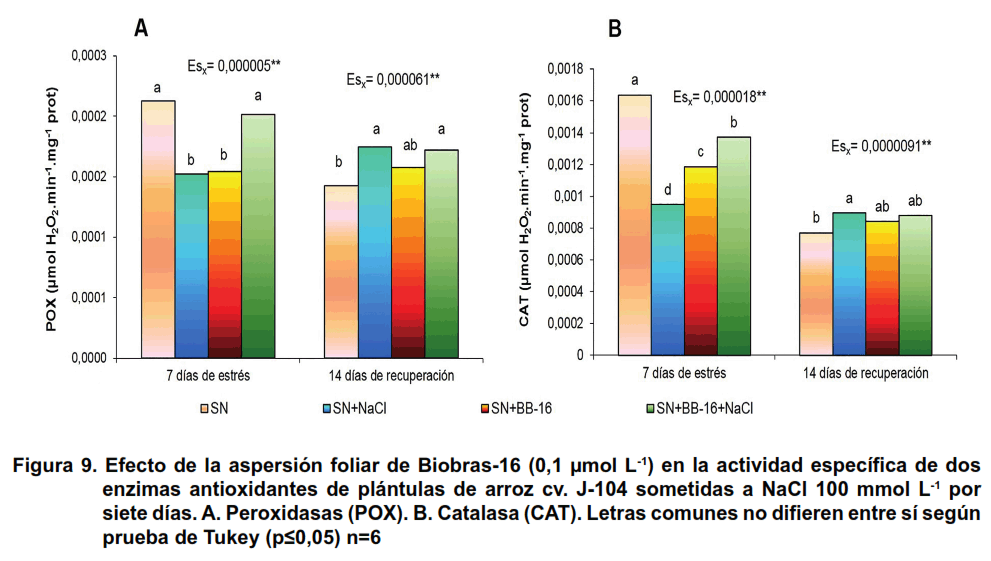
En la Figura 9 se observan las respuestas de las enzimas peroxidasa y catalasa de las plántulas de arroz cultivar J-104 sometidas a estrés salino. La actividad de ambas enzimas disminuyó significativamente al finalizar el tratamiento con NaCl. La aspersión foliar con BB-16 revirtió este comportamiento, incrementando la actividad específica de ambas enzimas (Figura 9A).
Catorce días después de la recuperación, se encontró que las plantas tratadas con NaCl presentaron un incremento significativo de las actividades de ambas enzimas y la aspersión foliar de BB-16 no modificó este comportamiento (Figura 9B).
La influencia del análogo BB-16 en el comportamiento de algunas enzimas antioxidantes de plántulas de arroz crecidas in vitro en presencia de NaCl 75 mM, mostraron que la exposición durante 16 días al BB-16 0,01 mg L-1 incrementó, significativamente, la actividad de las enzimas catalasa (CAT), superóxido dismutasa (SOD), glutatión reductasa y ligeramente la actividad de la ascorbato peroxidasa (3). Por otra parte, la aplicación de 1 µmol L-1 de DI-31, ingrediente activo del BB-16, a plantas de lechuga sometidas a tratamiento de NaCl 100 mmol L-1 durante cinco días fue capaz de revertir los efectos adversos que la salinidad provocó en estas plantas (38).
En frijol sometido a estrés salino, la aspersión foliar de EBL causó un incremento en la actividad SOD y no produjo modificaciones en la actividad POX en comparación con las plantas controles (22).Igualmente, se obtuvo un incremento en la SOD y la CAT por la aplicación exógena de brasinoesteroides en posturas de pepino sometidas a estrés salino (23). Por otra parte, en guisante, el tratamiento a las semillas con EBL provocó un incremento de la actividad de las enzimas CAT, POX y SOD en comparación con las plantas controles (39). Recientemente, se ha demostrado que, en arroz, la efectividad de la aplicación de la EBL en proteger a las plántulas ante el estrés de metales pesados (Cr) está asociada a la sobre-regulación de las enzimas antioxidantes (40).
CONCLUSIÓN
De forma general, se puede concluir que la aspersión foliar con Biobras-16 estimula el crecimiento de las plantas de arroz sometidas a estrés salino lo que puede estar asociada a un incremento de las actividades de las enzimas antioxidantes catalasa y peroxidasa al finalizar el período de estrés, igualmente pudiera asociarse al aumento en la concentración de clorofilas, fundamentalmente de la clorofila a.
NOTAS AL PIE
AOrellana, R.; Febles, J. y Ortega, F. GEO Cuba. Evaluación del medio ambiente cubano. Ed. Centenario SA, 2009, Santo Domingo, República Dominicana, 293 p.
BAlfonso, R. Determinación de parámetros genético-fisiológicos indicadores del estrés hídrico para su empleo en el mejoramiento genético del arroz (Oryza sativa L.) y la estabilidad varietal. Tesis de Doctorado, I.I.A., 1998.
CYoshida, S. Fundamentals of rice crop science. Ed. International Rice Research Institute, 1981, Los Baños, Philippines.
BIBLIOGRAFÍA
1. FAO. FAO Land and Plant Nutrition Management Service [en línea]. 2013, [Consultado: 7 de mayo de 2013], Disponible en: <http://www.fao.org/ag/agl/agll/spush>.
2. Yang, C.-J.; Zhang, C.; Lu, Y.-N.; Jin, J.-Q. y Wang, X.-L. “The Mechanisms of Brassinosteroids Action: From Signal Transduction to Plant Development”. Molecular Plant, vol. 4, no. 4, 2011, pp. 588-600, ISSN 1674-2052, DOI 10.1093/mp/ssr020.
3. Núñez, M. Brasinoesteroides y la respuesta de las plantas al estrés ambiental. Ed. Ediciones INCA, 2012, Mayabeque, Cuba, 80 p., ISBN 978-959-7023-57-9.
4. Yusuf, M.; Fariduddin, Q. y Ahmad, A. “24-Epibrassinolide modulates growth, nodulation, antioxidant system, and osmolyte in tolerant and sensitive varieties of Vigna radiata under different levels of nickel: A shotgun approach”. Plant Physiology and Biochemistry, vol. 57, 2012, pp. 143-153, ISSN 0981-9428, DOI 10.1016/j.plaphy.2012.05.004.
5. Núñez, V. M.; Reyes, G. Y.; Rosabal, A. L. y Martínez, G. L. “Análogos espirostánicos de brasinoesteroides y sus potencialidades de uso en la agricultura”. Cultivos Tropicales, vol. 35, no. 2, 2014, pp. 34-42, ISSN 0258-5936.
6. Fariduddin, Q.; Yusuf, M.; Ahmad, I. y Ahmad, A. “Brassinosteroids and their role in response of plants to abiotic stresses”. Biologia Plantarum, vol. 58, no. 1, 2014, pp. 9-17, ISSN 0006-3134, 1573-8264, DOI 10.1007/s10535-013-0374-5.
7. Núñez, V. M. de la C. Análogos de brasinoesteroides, nuevos productos para la agricultura [en línea]. Ed. Editorial Académica Española, 2013, Saarbrücken, 64 p., ISBN 978-3-659-08256-6, [Consultado: 18 de noviembre de 2016], Disponible en: <http://nbn-resolving.de/urn:nbn:de:101:1-201312147447>.
8. Hernández, J. A.; Pérez, J. J. M.; Bosch, I. D. y Castro, S. N. Clasificación de los suelos de Cuba 2015. Ed. Ediciones INCA, 2015, Mayabeque, Cuba, 93 p., ISBN 978-959-7023-77-7.
9. Paneque, P. V. M.; Calaña, N. J. M.; Calderón, V. M.; Borges, B. Y.; Hernández, G. T. C. y Caruncho, C. M. Manual de técnicas analíticas para análisis de suelo, foliar, abonos orgánicos y fertilizantes químicos [en línea]. Ed. Ediciones INCA, 2010, La Habana, Cuba, 157 p., ISBN 978-959-7023-51-7, [Consultado: 27 de enero de 2016], Disponible en: <http://mst.ama.cu/578/>.
10. Bates, L. S.; Waldren, R. P. y Teare, I. D. “Rapid determination of free proline for water-stress studies”. Plant and Soil, vol. 39, no. 1, 1973, pp. 205-207, ISSN 0032-079X, 1573-5036, DOI 10.1007/BF00018060.
11. Hodges, D. M.; DeLong, J. M.; Forney, C. F. y Prange, R. K. “Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds”. Planta, vol. 207, no. 4, 1999, pp. 604-611, ISSN 0032-0935, 1432-2048, DOI 10.1007/s004250050524.
12. Arnon, D. I. “Copper enzymes in isolated chloroplasts. polyphenoloxidase in Beta vulgaris”. Plant Physiology, vol. 24, no. 1, 1949, pp. 1-15, ISSN 0032-0889.
13. Bergmeyer, H. U. “Peroxidase” [en línea]. En: eds. Bergmeyer H. U. y Gawehn K., Methods of enzymatic analysis. Volume 4 Volume 4, Ed. Verlag Chemie, Weinheim, 1974, pp. 685-690, ISBN 978-0-323-16132-9, [Consultado: 18 de noviembre de 2016], Disponible en: <http://public.eblib.com/choice/publicfullrecord.aspx?p=1178409>.
14. Aebi, H. “Catalase in vitro”. Methods in Enzymology, vol. 105, 1984, pp. 121-126, ISSN 0076-6879, DOI 10.1016/S0076-6879(84)05016-3.
15. Bradford, M. M. “A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding”. Analytical Biochemistry, vol. 72, no. 1-2, 1976, pp. 248-254, ISSN 0003-2697, DOI 10.1016/0003-2697(76)90527-3.
16. Tukey, J. W. “Bias and confidence in not quite large samples”. The Annals of Mathematical Statistics, vol. 29, no. 2, junio de 1958, pp. 614-623, ISSN 0003-4851, DOI 10.1214/aoms/1177706647.
17. Statistical Graphics Crop. STATGRAPHICS® Plus [en línea]. (ser. Profesional), versión 5.1, [Windows], 2000, Disponible en: <http://www.statgraphics.com/statgraphics/statgraphics.nsf/pd/pdpricing>.
18. Rahdari, P. y Hoseini, S. M. “Salinity stress: a review”. Technical Journal of Engineering and Applied Sciences, vol. 1, no. 3, 2011, pp. 63–66, ISSN 2051-0853.
19. Morales, D.; Bolarín, M. del C. y Cayuela, E. “Respuesta de plantas de arroz (Oryza sativa L.) a la aplicación de diferentes niveles de NaCl. I. Crecimiento y relaciones hídricas”. Cultivos Tropicales, vol. 27, no. 4, 2006, pp. 27–33, ISSN 1819-4087.
20. Negrão, S.; Courtois, B.; Ahmadi, N.; Abreu, I.; Saibo, N. y Oliveira, M. M. “Recent Updates on Salinity Stress in Rice: From Physiological to Molecular Responses”. Critical Reviews in Plant Sciences, vol. 30, no. 4, 2011, pp. 329-377, ISSN 0735-2689, 1549-7836, DOI 10.1080/07352689.2011.587725.
21. Núñez, V. M.; Reyes, G. Y.; Rosabal, A. L.; Martínez, L. y González, C. M. C. “Brasinoesteroides y sus análogos estimulan el crecimiento de plántulas de dos genotipos de arroz (Oryza sativa L.) en medio salino”. Cultivos Tropicales, vol. 34, no. 1, 2013, pp. 74-80, ISSN 0258-5936.
22. Rady, M. M. “Effect of 24-epibrassinolide on growth, yield, antioxidant system and cadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmium stress”. Scientia Horticulturae, vol. 129, no. 2, 2011, pp. 232-237, ISSN 0304-4238, DOI 10.1016/j.scienta.2011.03.035.
23. Jiang, Y. P.; Cheng, F.; Zhou, Y. H.; Xia, X. J.; Mao, W. H.; Shi, K.; Chen, Z. X. y Yu, J. Q. “Brassinosteroid-induced CO2 assimilation is associated with increased stability of redox-sensitive photosynthetic enzymes in the chloroplasts in cucumber plants”. Biochemical and Biophysical Research Communications, vol. 426, no. 3, 2012, pp. 390-394, ISSN 0006-291X, DOI 10.1016/j.bbrc.2012.08.100.
24. Talaat, N. B. y Shawky, B. T. “24-Epibrassinolide alleviates salt-induced inhibition of productivity by increasing nutrients and compatible solutes accumulation and enhancing antioxidant system in wheat (Triticum aestivum L.)”. Acta Physiologiae Plantarum, vol. 35, no. 3, 2013, pp. 729-740, ISSN 0137-5881, 1861-1664, DOI 10.1007/s11738-012-1113-9.
25. Özdemir, F.; Bor, M.; Demiral, T. y Türkan, İ. “Effects of 24-epibrassinolide on seed germination, seedling growth, lipid peroxidation, proline content and antioxidative system of rice (Oryza sativa L.) under salinity stress”. Plant Growth Regulation, vol. 42, no. 3, 2004, pp. 203-211, ISSN 0167-6903, DOI 10.1023/B:GROW.0000026509.25995.13.
26. Ali, A. A. y Abdel-Fattah, R. I. “Osmolytes-antioxidant behaviour in Phaseolus vulgaris and Hordeum vulgare with brassinosteroid under salt stress”. Journal of Agronomy, vol. 5, no. 1, 2006, pp. 167–174, ISSN 1812-5379.
27. Siringam, K.; Juntawong, N.; Cha-um, S. y Kirdmanee, C. “Salt stress induced ion accumulation, ion homeostasis, membrane injury and sugar contents in salt-sensitive rice (Oryza sativa L. spp. indica) roots under isoosmotic conditions”. African Journal of Biotechnology, vol. 10, no. 8, 2011, pp. 1340-1346, ISSN 1684-5315.
28. Ekinci, M.; Yildirim, E. y Dursun, A. “Mitigation of Salt Stress in Lettuce (Lactuca sativa L. var. Crispa) by Seed and Foliar 24-epibrassinolide Treatments”. HortScience, vol. 47, no. 5, 2012, pp. 631-636, ISSN 0018-5345, 2327-9834.
29. Abd El-Samad, H. M.; Shaddad, M. A. K. y Barakat, N. “The role of amino acids in improvement in salt tolerance of crop plants”. Journal of Stress Physiology & Biochemistry, vol. 6, no. 3, 2010, pp. 25-37, ISSN 1997-0838.
30. Amini, F. y Ehsanpour, A. A. “Soluble proteins, proline, carbohydrates and Na+/K+ changes in two tomato (Lycopersicon esculentum Mill.) cultivars under in vitro salt stress”. American Journal of Biochemistry and Biotechnology, vol. 1, no. 4, 2005, pp. 212-216, ISSN 1553-3468.
31. Ben Rejeb, K.; Abdelly, C. y Savouré, A. “How reactive oxygen species and proline face stress together”. Plant Physiology and Biochemistry, vol. 80, 2014, pp. 278-284, ISSN 0981-9428, DOI 10.1016/j.plaphy.2014.04.007.
32. Hamilton, E. W. y Heckathorn, S. A. “Mitochondrial Adaptations to NaCl. Complex I Is Protected by Anti-Oxidants and Small Heat Shock Proteins, Whereas Complex II Is Protected by Proline and Betaine”. Plant Physiology, vol. 126, no. 3, 2001, pp. 1266-1274, ISSN 1532-2548, DOI 10.1104/pp.126.3.1266.
33. Nounjan, N. y Theerakulpisut, P. “Effects of exogenous proline and trehalose on physiological responses in rice seedlings during salt-stress and after recovery”. Plant Soil Environ, vol. 58, no. 7, 2012, pp. 309–315, ISSN 1805-9368.
34. Hossain, M. A. y Fujita, M. “Evidence for a role of exogenous glycinebetaine and proline in antioxidant defense and methylglyoxal detoxification systems in mung bean seedlings under salt stress”. Physiology and Molecular Biology of Plants, vol. 16, no. 1, 2010, pp. 19-29, ISSN 0971-5894, 0974-0430, DOI 10.1007/s12298-010-0003-0.
35. Ben Ahmed, C.; Ben Rouina, B.; Sensoy, S.; Boukhriss, M. y Ben Abdullah, F. “Exogenous Proline Effects on Photosynthetic Performance and Antioxidant Defense System of Young Olive Tree”. Journal of Agricultural and Food Chemistry, vol. 58, no. 7, 2010, pp. 4216-4222, ISSN 0021-8561, DOI 10.1021/jf9041479.
36. Zheng, C.; Jiang, D.; Liu, F.; Dai, T.; Jing, Q. y Cao, W. “Effects of salt and waterlogging stresses and their combination on leaf photosynthesis, chloroplast ATP synthesis, and antioxidant capacity in wheat”. Plant Science, vol. 176, no. 4, 2009, pp. 575-582, ISSN 0168-9452, DOI 10.1016/j.plantsci.2009.01.015.
37. Shahid, M. A.; Pervez, M. A.; Balal, R. M.; Ayyub, C. M.; Ghazanfar, U.; Abbas, T.; Rashid, A.; Garcia-Sanchez, F.; Mattson, N. S. y Akram, A. “Effect of salt stress on growth, gas exchange attributes and chlorophyll contents of pea (Pisum sativum)”. African Journal of Agricultural Research, vol. 6, no. 27, 2011, pp. 5808–5816, ISSN 1991-637X, DOI 10.5897/AJAR11.1087.
38. Serna, M.; Coll, Y.; Zapata, P. J.; Botella, M. Á.; Pretel, M. T. y Amorós, A. “A brassinosteroid analogue prevented the effect of salt stress on ethylene synthesis and polyamines in lettuce plants”. Scientia Horticulturae, vol. 185, 2015, pp. 105-112, ISSN 0304-4238, DOI 10.1016/j.scienta.2015.01.005.
39. Shahid, M. A.; Pervez, M. A.; Balal, R. M.; Mattson, N. S.; Rashid, A.; Ahmad, R.; Ayyub, C. M. y Abbas, T. “Brassinosteroid (24-epibrassinolide) enhances growth and alleviates the deleterious effects induced by salt stress in pea (Pisum sativum L.)”. Australian Journal of Crop Science, vol. 5, no. 5, 2011, pp. 500-510, ISSN 1835-2707.
40. Sharma, P.; Kumar, A. y Bhardwaj, R. “Plant steroidal hormone epibrassinolide regulate – Heavy metal stress tolerance in Oryza sativa L. by modulating antioxidant defense expression”. Environmental and Experimental Botany, vol. 122, 2016, pp. 1-9, ISSN 0098-8472, DOI 10.1016/j.envexpbot.2015.08.005.
Recibido: 09/12/2015
Aceptado: 29/04/2016
M.Cs. Yanelis Reyes Guerrero, Instituto Nacional de Ciencias Agrícolas. Gaveta Postal 1, San José de las Lajas, provincia Mayabeque, CP 32700. Email: yanelisrg@inca.edu.cu