Efecto de la morfología de los embriones somáticos en la regeneración de plantas de soya (Glycine max L. Merrill)
Effect of the somatic embryos morphology on plant regeneration of soybean (Glycine max L. Merrill)
Jorge L. Pérez-Pérez,I, II Lourdes García-Rodríguez,II Novisel Veitía-Rodríguez,II Idalmis Bermúdes-Caraballoso,II Raúl Collado-López,II Damaris Torres-RodríguezII
ICentro de Estudios de Biotecnología Vegetal. Universidad de Granma. Carretera a Manzanillo, km 17.5, Peralejo, Bayamo, Granma, Cuba.
IIInstituto de Biotecnología de las Plantas. Universidad Central “Marta Abreu” de Las Villas, Carretera a Camajuaní km 5.5, Santa Clara, Villa Clara, Cuba.
RESUMEN
Los protocolos establecidos para la regeneración de plantas vía embriogénesis somática en soya (Glycine max L. Merrill), son específicos para un número reducido de cultivares, lo que requiere de nuevos estudios que permitan disponer de protocolos más eficientes y reproducibles en un mayor número de genotipos. Este trabajo tuvo como objetivo determinar el efecto del tipo de morfología de los embriones somáticos en la germinación y conversión a planta en soya cultivar ʻIncasoy-27ʼ. Se emplearon embriones somáticos maduros con diferentes tipos de morfologías, que se colocaron en medio de cultivo de germinación. A los 30 días, se cuantificó el número de embriones somáticos con germinación completa o parcial por tipo de morfología. En la fase de conversión se determinó a los diez días de cultivo, el porcentaje de supervivencia de las plantas regeneradas y se realizó una caracterización morfológica de las plantas regeneradas en comparación con las plantas obtenidas de semilla botánica. Se logró un 74,38 % de embriones somáticos germinados con morfología normal y anormal. El porcentaje de supervivencia de las plantas regeneradas alcanzó el 70,0 % de eficiencia. La descendencia obtenida a partir de las plantas regeneradas de embriones somáticos, fueron morfológicamente similares a las plantas control. Los resultados derivados de esta investigación permitieron demostrar que es posible la regeneración de plantas completas vía embriogénesis somática en soya cultivar cubano ʻIncasoy-27ʼ.
Palabras clave: cultivo de tejidos, embriogénesis somática, medio de cultivo.
ABSTRACT
The established protocols for plant regeneration via somatic embryogenesis in soybean (Glycine max L. Merrill) are specific to a small number of cultivar, requiring new studies to provide more efficient and reproducible protocols in a large number of genotypes. This work had as objective to determine the effect of the morphology type of somatic embryos on the germination and plant conversion in soybean cultivar ʻIncasoy-27ʼ. Mature somatic embryos were used with different morphologies types that were placed in germination medium. At 30 days the somatic embryos number with complete or partial germination by morphology type was quantified. In the conversion phase it was determined to the 10 days of cultivation the survival percentage of the regenerated plants and it was carried out a morphological characterization of the regenerated plants in comparison with the obtained plants of seeds. It was achieved at 74,38 % of somatic embryos germinated with normal and abnormal morphology. The percentage of survival of regenerated plants reached 70,0 % efficiency. The progeny obtained from the regenerated plants of somatic embryos were morphologically similar to the control plants. The results from this research show that it is possible to allow regeneration of whole plants through somatic embryogenesis in soybean Cuban cultivar ʻIncasoy-27ʼ.
Key words: tissue culture, somatic embryogenesis, culture medium.
INTRODUCCIÓN
La embriogénesis somática es un proceso inductivo que permite que células vegetales no cigóticas, formen embriones somáticos con una estructura bipolar y sin conexión vascular con el tejido original (1, 2). Además, debido a la no fusión de los gametos es posible lograr una alta homogeneidad genética en las plantas regeneradas (3).
Esta vía de regeneración de plantas ha sido implementada de manera eficiente en diferentes especies; sin embargo, en soya se ha descrito en un número limitado de genotipos (4), entre los que destacan los de origen norteamericano, Jack, Williams y Fayette así como los brasileños Bragg y IAS5 (5). Para ello, se requiere del empleo de reguladores del crecimiento como las auxinas, que estimulan las células con capacidad embriogénica e inducen la formación de los embriones somáticos (1). No obstante, existen evidencias que demuestran que dentro de las auxinas, el papel protagónico le corresponde al ácido 2,4-diclorofenoxiacético (2,4-D). Este regulador del crecimiento, permite inducir una embriogénesis somática repetitiva, donde la formación de los embriones somáticos secundarios tiene un origen unicelular y ocurre a partir de embriones somáticos primarios (6).
Algunas referencias asocian la formación de una célula embriogénica, con el 2,4-D y los cambios que ocurren en la estructura de la cromatina por metilación del ácido desoxirribonucleico (ADN), que conducen a una reprogramación genómica en las células somáticas y se expresan los genes requeridos para la expresión embriogénica (7). Sin embargo, el uso prolongado de altas concentraciones de 2,4-D en los medios de cultivo de inducción y multiplicación de embriones somáticos en soya, puede conllevar a la aparición de diversos tipos de morfologías anormales durante la fase de maduración, que afecta la eficiencia de la germinación y conversión a planta (8).
Algunos autores han encontrado aberraciones e inestabilidad cromosómica en embriones somáticos de soya, atribuido a los reguladores de crecimiento durante el cultivo in vitro (9). Debido a esto la mayoría de los investigadores desechan los embriones somáticos de soya que muestran morfologías anormales, por lo que son escasas las referencias que abordan la eficiencia de germinación por tipo de morfología. Otros no emplean reguladores del crecimiento en la maduración y germinación de embriones somáticos; o mediante la manipulación de las condiciones de cultivo, como es el caso de la desecación parcial, se logra prevenir la germinación precoz (10).
Teniendo en cuenta que el empleo de altas concentraciones de 2,4-D puede causar diferentes morfologías anormales, se realizó este trabajo con el objetivo de determinar el efecto del tipo de morfología de los embriones somáticos en la eficiencia de germinación y conversión a planta en soya cultivar ʻIncasoy-27ʼ.
MATERIALES Y MÉTODOS
El trabajo se realizó en el Instituto de Biotecnología de las Plantas en Santa Clara, Cuba. Se empleó el cultivar cubano de soya ʻIncasoy-27ʼ, obtenido en el Instituto Nacional de Ciencias Agrícolas (INCA) de Cuba, mediante hibridación natural, que ocurrió en el genotipo brasileño ‘BR-32’.
Condiciones de cultivo
El experimento se desarrolló en cámara de crecimiento con luz solar, con una duración del período luminoso de 12 a 13 horas de luz; temperatura de 26±1 °C; intensidad del flujo de fotones fotosintéticos entre 68,0 a 72,0 μmol m-2s-1, medido con un luxómetro (Extech 401025, EUA).
Se tomaron embriones somáticos con dos meses en medio de cultivo de maduración (10), que contenía maltosa (6,0 %) como fuente de carbono. Para la desecación parcial de los embriones somáticos, estos se colocaron durante cuatro días en placas de Petri (100 × 10 mm) estériles. Cada placa contenía alrededor de 48 embriones somáticos que se colocaron hacia un extremo de la misma y en el lado opuesto un fragmento de medio de cultivo semisólido (1,0 cm3), según la metodología de Bailey (11).
Germinación de los embriones somáticos
Se colocaron de 11 a 12 embriones somáticos maduros (repeticiones) con desecación parcial por frasco de vidrio de 250 mL de capacidad, distribuidos en 21 frascos para un total de 242 embriones somáticos. Cada frasco de cultivo contenía 30 mL de medio de cultivo de germinación, con las sales MS (12), vitaminas B5 (13), sacarosa 1,5 %, Gelrite ® 0,3 % y pH 5,8. A los 30 días de cultivo se cuantificó el total de embriones somáticos germinados y en base a este valor se determinó el número de embriones somáticos con germinación completa o parcial (%) por tipo de morfología.
Conversión a planta
Posteriormente, fueron seleccionadas 100 plantas in vitro procedentes del medio de cultivo de germinación. Para lograr homogeneidad entre las plantas se tomaron aquellas que tenían entre 5,0-6,0 cm longitud y subcultivadas a medio de cultivo de germinación durante cinco días. Los frascos se taparon con papel de aluminio y para propiciar una mayor ventilación, se realizó un orificio en la tapa con ayuda de un lápiz, en el primer y tercer día de cultivo. Al finalizar las raíces se lavaron con agua destilada estéril, antes de transferir las plantas a condiciones de invernadero. La esterilización del agua se realizó en autoclave a 121 °C y 1,1 kg cm-2 de presión durante 40 minutos.
La aclimatización de las plantas en condiciones de cultivo ex vitro se realizó en el período de invierno (marzo–abril). Las plantas se colocaron en frascos de policarbamato (500 mL) que contenían de forma homogénea, un sustrato estéril con 75 % de materia orgánica descompuesta de origen vacuno y un 25 % del mineral zeolita. Este último es un alumino-silicato natural cristalino microporoso, con excelentes propiedades de intercambio iónico, alto poder de absorción y granulación 1,0-3,0 mm (14). La esterilización del sustrato se realizó previamente en estufa a 180 °C durante dos horas.
Las plantas control se obtuvieron a partir de semillas botánicas en iguales condiciones de cultivo, con dos riegos diarios por microaspersión de tres minutos de duración; temperatura media de 27 ±2 °C y humedad relativa 84 ±5 %. La luz solar se reguló con una malla sombreadora de color negro, a una intensidad del flujo de fotones fotosintéticos de 280 a 400 μmol m-2s-1.
A los diez días se determinó la eficiencia de conversión, a partir del número de plantas vivas (%), y a los 30 días se evaluaron los caracteres morfológicos de las plantas regeneradas procedentes del cultivo in vitro. De igual forma fueron evaluadas a los 75 días de cultivo, las plantas controles y la descendencia de plantas regeneradas in vitro, que fueron obtenidas a partir de semillas maduras.
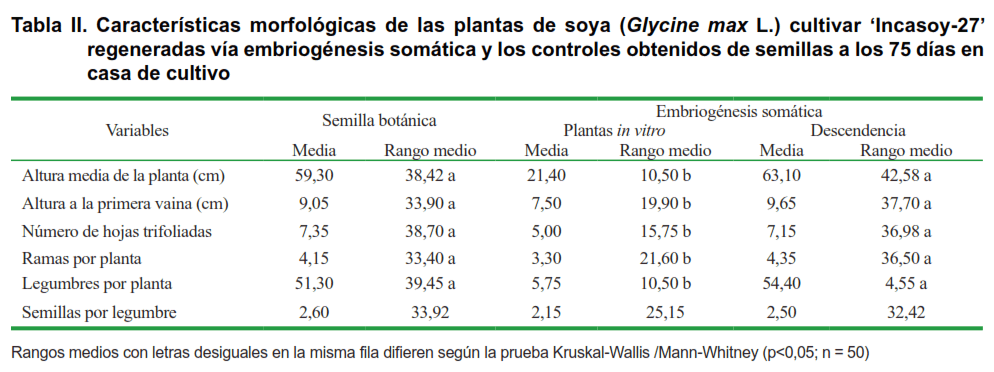
Para la evaluación morfológica se seleccionaron 50 plantas al azar, de cada uno de los tres tratamientos (plantas obtenidas de semilla botánica, plantas regeneradas a partir de embriones somáticos, y descendencia de las plantas procedentes del cultivo de tejidos) y para su comparación se utilizó el descriptor de este cultivar. Los criterios de evaluación fueron: color de las hojas; número de foliolos en las hojas; color de la flor; altura media de la planta (cm); altura a la primera vaina (cm); número de hojas por planta; legumbres por planta; granos por legumbre y color de la semilla. Para la evaluación visual de estos caracteres, se tuvo en cuenta el descriptor morfológico de este genotipo. La altura de las plantas se determinó con una regla milimetrada y se midió desde la base del tallo hasta el meristemo apical de la planta.
Análisis estadístico
Para el desarrollo de la investigación se utilizó un diseño completamente al azar con cinco repeticiones en el tiempo. Los datos se analizaron mediante la prueba de Kruskal-Wallis (15)y las diferencias entre tratamientos con la prueba de Mann Whitney. Se empleó el programa estadístico SPSS (del inglés: Statistic Package for Social Science) versión PASW Statistics 18, con un valor de p<0,05 (16).
RESULTADOS Y DISCUSIÓN
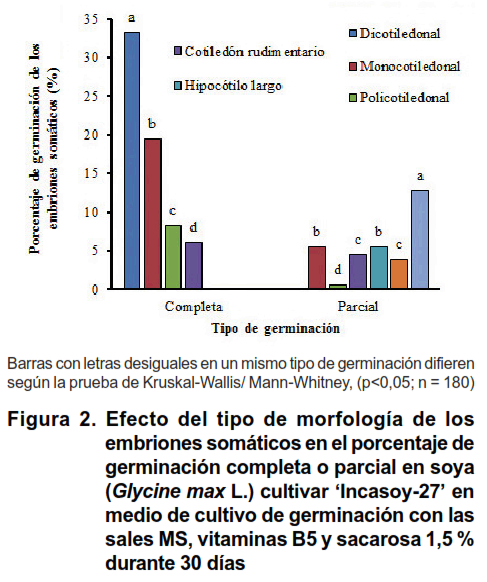
La evaluación de la morfología de los embriones somáticos demostró que esta influyó en su capacidad para germinar y regenerar plantas de soya cultivar ʻIncasoy-27ʼ. Los embriones somáticos que tenían una morfología de tipo dicotiledonal, desarrollaron una germinación completa donde eran visibles los primordios foliares simples de color verde; con alargamiento en las raíces, hipocótilo y epicótilo (Figura 1A, B). Por el contrario, los embriones somáticos con morfologías de tipo hipocótilo largo y fusionados tenían germinación parcial, los cuales solo desarrollaron la raíz (Figura 1C, D). No obstante, se encontraron embriones somáticos principalmente de tipo fusionados, que no lograron germinar al no desarrollar ningún meristemo (Figura 1E).
Aunque se logró la germinación en los embriones somáticos maduros, se encontró que los que tenían morfologías de tipo anormal representaban el mayor número, con un valor equivalente al 49,59 % distribuidos de la siguiente manera: monocotiledonal (18,60 %),policotiledonal (6,61 %), cotiledón rudimentario (7,85 %), hipocótilo largo (4,13 %), fasciado (2,89 %) y fusionados (9,51 %). Sin embargo, el número de embriones somáticos con morfologías anormales, duplicó el valor obtenido en los embriones somáticos normales, con diferencias significativas entre ellos (Tabla I).
Aunque fue posible la germinación de embriones somáticos con morfologías anormales, existen algunos tipos como trompeta, que tienen un limitado desarrollo de meristemo caulinar y otros del tipo fasciados que logran formar el meristemo radical, pero nunca el meristemo caulinar, lo que afecta su capacidad para germinar (17).
Estos cambios en la simetría bilateral de los embriones somáticos, puede deberse a factores endógenos como los inhibidores del transporte polar de las auxinas durante el desarrollo del embrión somático; así como, por la continua exposición del tejido a las altas concentraciones de 2,4-D, que interfiere con el gradiente de auxina polar que se establece en la embriogénesis somática (18, 19).
No obstante, del total de embriones somáticos germinados los de tipo dicotiledonal presentaron en su totalidad germinación completa (Figura 2). Los embriones somáticos con otros tipos de morfologías clasificadas como anormales, presentaron germinación completa y parcial. Del total de embriones somáticos con germinación completa (67,21 %), los que tenían morfología dicotiledonal tuvieron el mayor porcentaje (33,33 %), seguido de los monocotiledonal, policotiledonal y cotiledón rudimentario, con diferencias significativas entre ellos (Figura 2). En los embriones somáticos anormales con germinación parcial (32,79 %), los de morfologías del tipo fusionados presentaron los mayores valores de germinación (12,78 %), seguido de monocotiledonal e hipocótilo largo (Figura 2).
Esta reducción en la capacidad de germinación de los embriones somáticos, también pudiera ser debido a una ineficiente acumulación de sustancias de reservas durante la fase de maduración, que limita su capacidad para recuperarse después del estrés provocado por el proceso de desecación.
Existen referencias que demuestran que la incorporación de ácido abscísico (ABA) durante las primeras etapas de desarrollo de la embriogénesis somática, favorece la síntesis de compuestos de reserva en los embriones somáticos (20). Este efecto del ABA se le atribuye a que regula diferentes procesos fisiológicos y activa genes vinculados con la respuesta al estrés (21); mientras que, un bajo contenido endógeno puede reducir la capacidad de germinación y afectar su recuperación posterior al proceso de desecación.
En otros estudios se ha encontrado mayor acumulación de sustancias de reserva en los embriones somáticos, en las primeras tres semanas, en medio de cultivo de maduración, pero al finalizar este período el contenido de los ácidos linoléico y linolénico declinaron, mientras que los ácidos palmítico y esteárico aumentaron. Esto pudo estar relacionado con el proceso de germinación de los embriones somáticos, atribuido a una removilización o degradación de algunas sustancias de reserva durante la senescencia, similar a lo que ocurre en otros tejidos vegetales (22).
La baja capacidad de germinación de los embriones somáticos, también se ha descrito en la literatura científica. En cultivares hindúes de soya después de un período de cinco días de desecación, el 40,9 % de los embriones somáticos lograron germinar (6). Los autores señalaron que las fases de maduración y germinación de embriones somáticos en soya, son poco eficiente en muchos estudios, debido al alto número de embriones somáticos con morfologías anormales que no logran formar plantas completas.
Según estudios realizados (4), después de siete días de desecación los embriones somáticos fueron colocados en dos concentraciones de sacarosa (1,5 y 3,0 %) y encontraron el mayor porcentaje de plantas con un crecimiento normal (53,0 %) en el genotipo indú ʻPS1477ʼ con 1,5 % de sacarosa.Este resultado es inferior al 67,21 % obtenido en el presente trabajo con igual concentración de sacarosa pero con cuatro días de desecación. Es posible que el empleo de siete días haya provocado una mayor deshidratación en los embriones somáticos y afectó su potencial de germinación.
A partir de los embriones somáticos con germinación completa en el cultivar ʻIncasoy-27ʼ , se obtuvieron plantas que lograron adaptarse a las condiciones de cultivo ex vitro. El porcentaje de supervivencia de las plantas regeneradas fue del 76,0 % de eficiencia de conversión a los 15 días de cultivo; mientras que un 100 % se logró en las plantas control obtenidas de semilla botánica.
Las plantas del tratamiento control (Figura 3A) y las regeneradas a partir de embriones somáticos, tenían características comunes como hojas trifoliadas de color verde y flores blancas (Figura 3B, C). Sin embargo, las plantas obtenidas mediante cultivo in vitro desarrollaron un tallo delgado y, en algunos casos, un limitado crecimiento y desarrollo foliar, atribuido a una habituación del tejido a las condiciones del cultivo in vitro (Figura 3C).
Las plantas regeneradas in vitro comenzaron a florecer y fructificar prematuramente alrededor de los 15 días (Figura 3D) a diferencia de las plantas controles, donde la floración ocurrió alrededor de los 35 a 40 días. Esto podría deberse a que las plantas in vitro provenían de un período de 45 días en fase de germinación que permitió desarrollar el sistema radical y apical. Por el contrario, las plantas de semillas botánicas necesitaron entre cinco y siete días para iniciar la germinación. Las semillas obtenidas de las plantas regeneradas in vitro, dieron origen a la descendencia con un crecimiento normal y las características propias de este genotipo (Figura 3E, F); en ningún caso, fueron detectadas plantas fuera de tipo.
Las plantas procedentes de embriones somáticos tuvieron un menor desarrollo que las plantas del tratamiento control. De igual forma no se encontró diferencias visibles en el color de las semillas, las que mostraron el color amarillo verdoso propio de este cultivar. En cuanto a los resultados obtenidos en la descendencia, estas plantas no tuvieron diferencias significativas con respecto a los valores obtenidos en los controles en iguales condiciones de cultivo (Tabla II).
La eficiencia de conversión obtenida fue superior a los valores alcanzados en los cultivares ʻIAS5ʼ y ʻBRSMG68ʼ (5). Estos autores obtuvieron un 45,0 % de conversión en el cultivar ʻBRSMG68ʼ y 42,0 % en ʻIAS5ʼ , atribuido a una germinación precoz de los embriones somáticos sin haber alcanzado la suficiente madurez, lo que provoca un pobre desarrollo en las plantas in vitro y reduce la eficiencia de regeneración. Esto también indica que la eficiencia de conversión a planta, pudiera estar influenciado por el genotipo.
Al respecto, en el cultivar brasileño de soya ʻCD 201ʼ, se logró un 82,0 % de conversión a planta (23). Sin embargo, esta eficiencia disminuyó en los cultivares ʻCD 220ʼ (58,0 %) y ʻCD 216ʼ (51,0 %), atribuido a la presencia de embriones somáticos con morfologías anormales, que lograron germinar pero no convirtieron a planta. Otros autores al concluir los 28 días de germinación (4), pasaron las plantas in vitro a frascos de cultivo que contenían una mezcla de los sustratos turba-perlita-vermiculita (2:1:1), que fue hidratada con las sales MS (25,0 %) y cultivadas 28 días a una temperatura de 26±1 °C y un fotoperíodo de 16 horas de luz (150 μmol m-2s-1). Durante ese período las plantas fueron endurecidas en condiciones controladas antes de pasar al invernadero, y permitió que después de 21 días de cultivo ex vitro lograran un 80,0 % de supervivencia en el cultivar ʻDS 2706ʼ.
En cuanto a las características de las plantas regeneradas, una de las causas que pudo haber provocado que las procedentes de embriones somáticos fueran más pequeñas que las obtenidas de semillas, es que las reservas de proteínas y lípidos en el embrión somático pueden declinar antes de ocurrir la elongación de la raíz, debido a la falta del tejido nutritivo que rodea la semilla (24). Esto difiere de las plantas que se obtienen a partir de semillas, donde las sustancias nutritivas se movilizan desde diferentes partes de la planta, hacia las semillas que están en desarrollo y luego estas sustancias contenidas en las semillas favorecen la germinación y desarrollo del brote.
CONCLUSIONES
La eficiencia de germinación de los embriones somáticos y su capacidad para convertir a planta, está relacionada con el tipo de morfología que estos desarrollen.
Los embriones somáticos con morfología de tipo dicotiledonal o normal presentaron en su totalidad germinación completa.
Los embriones somáticos con morfologías anormales como monocotiledonal, policotiledonal y cotiledón rudimentario, tienen germinación completa y parcial.
El porcentaje de supervivencia de las plantas regeneradas a partir de embriones somáticos, está influenciado por el tipo de morfología y la eficacia en la maduración de los embriones somáticos.
BIBLIOGRAFÍA
1. Joshi, R. y Kumar, P. “Regulation of somatic embryogenesis in crops: a review”. Agricultural Reviews, vol. 34, no. 1, 2013, pp. 1-20, ISSN 0253-1496.
2. Fehér, A. “Somatic embryogenesis - Stress-induced remodeling of plant cell fate”. Biochimica Et Biophysica Acta, vol. 1849, no. 4, 2015, pp. 385-402, ISSN 0006-3002, DOI 10.1016/j.bbagrm.2014.07.005.
3. Smertenko, A. y Bozhkov, P. “The Life and Death Signalling Underlying Cell Fate Determination During Somatic Embryogenesis” [en línea]. En: eds. Nick P. y Opatrny Z., Applied Plant Cell Biology, Ed. Springer Berlin Heidelberg, Berlin, Heidelberg, 2014, pp. 131-178, ISBN 978-3-642-41786-3, DOI 10.1007/978-3-642-41787-0_5, [Consultado: 12 de febrero de 2017], Disponible en: <http://link.springer.com/10.1007/978-3-642-41787-0_5>.
4. Huynh, H. N.; Lal, S. K.; Singh, S. K.; Prabhu, K. V. y Talukdar, A. “High frequency regeneration in soybean (Glycine max (L.) Merrill.) through direct somatic embryogenesis from immature cotyledons”. Indian Journal of Plant Physiology, vol. 20, no. 3, 2015, pp. 232-239, ISSN 0019-5502, 0974-0252,
DOI 10.1007/s40502-015-0166-3.
5. Droste, A.; da Silva, A. M.; de Souza, I. F.; Wiebke-Strohm, B.; Bücker-Neto, L.; Bencke, M.; Sauner, M. V. y Bodanese-Zanettini, M. H. “Screening of Brazilian soybean genotypes with high potential for somatic embryogenesis and plant regeneration”. Pesquisa Agropecuária Brasileira, vol. 45, no. 7, 2010, pp. 715-720, ISSN 0100-204X, DOI 10.1590/S0100-204X2010000700012.
6. Mariashibu, T. S.; Subramanyam, K.; Arun, M.; Theboral, J.; Rajesh, M.; Rengan, S. K.; Chakravarthy, R.; Manickavasagam, M. y Ganapathi, A. “Assessment of somatic embryogenesis potency in Indian soybean (Glycine max (L.) Merr.) cultivars”. Indian Journal of Experimental Biology, vol. 51, no. 10, 2013, pp. 849-859, ISSN 0975-1009, 0019-5189.
7. Us-Camas, R.; Rivera-Solís, G.; Duarte-Aké, F. y De-la-Peña, C. “In vitro culture: an epigenetic challenge for plants”. Plant Cell, Tissue and Organ Culture (PCTOC), vol. 118, no. 2, 2014, pp. 187-201, ISSN 0167-6857, 1573-5044, DOI 10.1007/s11240-014-0482-8.
8. Verma, K.; Saini, R. y Rani, A. “Recent advances in the regeneration and genetic transformation of soybean”. Journal of Innovative Biology March, vol. 1, no. 1, 2014, pp. 015–026, ISSN 2348-6473.
9. Homrich, M. S.; Wiebke-Strohm, B.; Weber, R. L. M. y Bodanese-Zanettini, M. H. “Soybean genetic transformation: a valuable tool for the functional study of genes and the production of agronomically improved plants”. Genetics and Molecular Biology, vol. 35, no. 4, 2012, pp. 998-1010, ISSN 1415-4757,
DOI 10.1590/S1415-47572012000600015.
10. Wiebke-Strohm, B.; Shenkel, M.; Mayer, R. L.; Droste, A. y Bodanese-Zanettini, M. H. “Strategies for improvement of soybean regeneration via somatic embryogenesis and genetic transformation”. En: ed. Barrera-Saldaña H. A., Genetic engineering: basics, new applications and responsibilities, Ed. InTech, Rijeka, 2011, pp. 145-172, ISBN 978-953-307-790-1.
11. Bailey, M. A.; Boerma, H. R. y Parrott, W. A. “Genotype-specific optimization of plant regeneration from somatic embryos of soybean”. Plant Science, vol. 93, no. 1-2, 1993, pp. 117–120, ISSN 0168-9452, DOI 10.1016/0168-9452(93)90040-7.
12. Murashige, T. y Skoog, F. “A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures”. Physiologia Plantarum, vol. 15, no. 3, 1962, pp. 473-497, ISSN 1399-3054, DOI 10.1111/j.1399-3054.1962.tb08052.x.
13. Gamborg, O. L.; Miller, R. A. y Ojima, K. “Nutrient requirements of suspension cultures of soybean root cells”. Experimental Cell Research, vol. 50, no. 1, 1968, pp. 151-158, ISSN 0014-4827,
DOI 10.1016/0014-4827(68)90403-5.
14. Posada, L.; Padrón, Y.; González, J.; Rodríguez, R.; Barbón, R.; Norman, O.; Rodríguez, R. C. y Gómez-Kosky, R. “Efecto del Pectimorf ® en el enraizamiento y la aclimatización in vitro de brotes de papaya (Carica papaya L.) cultivar Maradol Roja”. Cultivos Tropicales, vol. 37, no. 3, 2016, pp. 50-59, ISSN 0258-5936, DOI 10.13140/RG.2.1.1642.2642.
15. Kruskal, W. H. y Wallis, W. A. “Use of Ranks in One-Criterion Variance Analysis”. Journal of the American Statistical Association, vol. 47, no. 260, 1952, pp. 583-621, ISSN 0162-1459, DOI 10.1080/01621459.1952.10483441.
16. IBM Corporation. PASW Statistics [en línea]. versión 18.0, [Windows], Multiplataforma, Ed. IBM Corporation, 2009, U.S, Disponible en: <http://www.ibm.com>.
17. Fernando, J. A.; Vieira, M. L. C.; Geraldi, I. O. y Appezzato-da-Glória, B. “Anatomical Study of Somatic Embryogenesis in Glycine max (L.) Merrill”. Brazilian Archives of Biology and Technology,
vol. 45, no. 3, 2002, pp. 277-286, ISSN 1516-8913, DOI 10.1590/S1516-89132002000300005.
18. Robert, H. S.; Grones, P.; Stepanova, A. N.; Robles, L. M.; Lokerse, A. S.; Alonso, J. M.; Weijers, D. y Friml, J. “Local Auxin Sources Orient the Apical-Basal Axis in Arabidopsis Embryos”. Current Biology,
vol. 23, no. 24, 2013, pp. 2506-2512, ISSN 0960-9822, DOI 10.1016/j.cub.2013.09.039.
19. Robert, H. S.; Grunewald, W.; Sauer, M.; Cannoot, B.; Soriano, M.; Swarup, R.; Weijers, D.; Bennett, M.; Boutilier, K. y Friml, J. “Plant embryogenesis requires AUX/LAX-mediated auxin influx”. Development, vol. 142, no. 4, 2015, pp. 702-711, ISSN 0950-1991, 1477-9129, DOI 10.1242/dev.115832.
20. Normah, M. N.; Rohani, E. R. y Mohamed-Hussein, Z. A. “Somatic embryogenesis in higher plants”. Malaysian Applied Biology, vol. 42, no. 2, 2013, pp. 1-12, ISSN 0126-8643.
21. Gomez-Cadenas, A.; Vives, V.; Zandalinas, S.; Manzi, M.; Sanchez-Perez, A.; Perez-Clemente, R. y Arbona, V. “Abscisic Acid: A Versatile Phytohormone in Plant Signaling and Beyond”. Current Protein and Peptide Science, vol. 16, no. 5, 2015, pp. 413-434, ISSN 1875-5550, DOI 10.2174/13892037166661503 30130102.
22. He, Y.; Young, T. E.; Clark, K. R.; Kleppinger-Sparace, K. F.; Bridges, W. C. y Sparace, S. A. “Developmental profile of storage reserve accumulation in soybean somatic embryos”. In Vitro Cellular & Developmental Biology - Plant, vol. 47, no. 6, 2011, pp. 725-733, ISSN 1054-5476, 1475-2689, DOI 10.1007/s11627-011-9375-2.
23. Texeira, L. R.; Braccini, A. de L. e; Churata, B. G. M.; Vieira, E. S. N.; Martins, P. K. y Schuster, I. “Evaluation of soybean cultivars on the embryogenic and organogenic potential”. Acta Scientiarum. Agronomy, vol. 33, no. 1, 2011, pp. 67-74, ISSN 1807-8621, DOI 10.4025/actasciagron.v33i1.12344.
24. Freire-Seijo, M. “Aspectos básicos de la embriogénesis somática”. Biotecnología Vegetal, vol. 3, no. 4, 2003, pp. 195-209, ISSN 2074-8647.
Recibido: 14/07/2016
Aceptado: 14/11/2016
Jorge L. Pérez Pérez, Centro de Estudios de Biotecnología Vegetal. Universidad de Granma. Carretera a Manzanillo, km 17.5, Peralejo, Bayamo, Granma, Cuba. Email. jperez@udg.co.cu