ARTÍCULO ORIGINAL
Acumulación de iones y tolerancia a la salinidad en diferentes cultivares cubanos de garbanzo (Cicer arietinum L.)
Ion accumulation and tolerance to salinity in differentcuban chickpea cultivars (Cicer arietinum L.)
Yanitza Meriño-Hernández,I Pedro Rodríguez-Hernández,II Omar Cartaya-Rubio,III José M. Dell'Amico-Rodríguez,III Tony Boicet-Fabré,I Tomás Shagarodsky-Scull,IV Yenisei Hernández-BarandaIII
I Departamento de Producción Agrícola. Universidad de Granma, carretera a Manzanillo km 17.5, Bayamo, Granma. Código Postal 85 100
II Corporación Colombiana de Investigación Agropecuaria (CORPOICA), Centro de Investigación Obonuco, . km 5, Vía Pasto-Obonuco, San Juan de Pasto, Nariño, Colombia
III Instituto Nacional de Ciencias Agrícolas (INCA), carretera San José-Tapaste, km 3½, Gaveta Postal 1, San José de Las Lajas, Mayabeque, Cuba. CP 32 700
IV Instituto de Investigaciones Fundamentales de Agricultura Tropical (INIFAT), calle 1 y 2, Santiago de las Vegas, Boyeros, Ciudad de la Habana, Cuba. CP 17 200
Resumen
El garbanzo (Cicer arietinum L.) es considerado una especie sensible a la salinidad, pero existen diferencias en cuanto al grado de tolerancia a este tipo de estrés. El presente estudio se realizó para determinar la tolerancia a la salinidad de algunos cultivares cubanos de garbanzo y su relación con indicadores del desarrollo y la acumulación de iones en los diferentes órganos de la planta. Para ello, se estudiaron ocho cultivares de garbanzo expuestos a dos tratamientos, 0 y 50 mM de NaCl. Los resultados evidenciaron que los cultivares N-29, Nac-5HA y JP-94 fueron los que mostraron mayor grado de tolerancia a la salinidad. El cultivar N-29 resultó ser más tolerante y aunque disminuyó su crecimiento, los resultados evidencian que también acumuló menos cantidad de iones en los diferentes órganos de la planta, en las dos condiciones de salinidad impuesta. Por tanto, se infiere que la reducción del crecimiento estuvo relacionado con alguna incapacidad que tienen las plantas de garbanzo para evitar que altas concentraciones de iones salinos lleguen a las hojas, trayendo como consecuencia una considerable variabilidad en cuanto al grado de tolerancia a la salinidad de los cultivares evaluados.
Palabras clave: biomasa, crecimiento, toxicidad, tolerantes.
ABSTRACT
Chickpea (Cicer arietinum L.) is considered a species sensitive to salinity, but there are differences in the degree of tolerance to this type of stress. The present study was carried out to determine the tolerance to salinity of some cuban cultivars of chickpea and its relationship with indicators of the development and accumulation of ions in the different organs of the plant. For this, eight chickpea cultivars exposed to two treatments, 0 and 50 mM NaCl, were studied. The results showed that the cultivars Nac-29, Nac-5HA and JP-94 were the ones that showed the highest degree of tolerance to salinity. The cultivar N-29 turned out to be more tolerant and although it diminished its growth, the results show that it also accumulated less quantity of ions in the different organs of the plant, in the two conditions of imposed salinity. Therefore, it can be inferred that the reduction in growth was related to some inability of the chickpea plants to prevent high concentrations of saline ions from reaching the leaves, resulting in considerable variability in the degree of tolerance to salinity the cultivars evaluated.
Key words: biomass, growth, toxicity, tolerant.
INTRODUCCIÓN
El estrés abiótico, en particular la sequía y la salinidad, no solo compromete la calidad de las cosechas y limita el rendimiento, sino también restringe las áreas de siembra y la viabilidad de la producción de las cosechas (1). Estos problemas ambientales en forma de estrés abiótico son básicamente una severa amenaza para la agricultura mundial (2).
Actualmente, la salinidad es un serio problema y está aumentando fuertemente en muchas partes del mundo, particularmente en las regiones áridas y semi áridas (3). Más del 74 % de los suelos dedicados a la agricultura a nivel mundial, presentan problemas de salinidad (4) y en muchos de los casos la conductividad eléctrica del suelo supera el índice de tolerancia de importantes especies económicas (4,5).
El exceso de sales en el suelo, provoca daños en las plantas que están relacionados con una toxicidad específica, normalmente asociada a la absorción excesiva de Na+ y Cl-, un desequilibrio nutricional provocado por la interferencia de los iones salinos con los nutrientes esenciales, un estrés hídrico que se produce por la disminución del potencial osmótico del medio y la combinación de los efectos antes indicados (6).
Diversos métodos han sido utilizados para disminuir el efecto perjudicial de las sales de los suelos y aumentar su calidad agroproductiva (7–9). Las tendencias actuales incluyen métodos biológicos y de manejo agrotécnico; entre ellos, la introducción y la selección de especies y variedades que presenten niveles de tolerancia, adaptabilidad y producción adecuados, que permitan sustituir las más susceptibles (9–11).
El garbanzo (Cicer arietinum L.) es una leguminosa de importancia comercial y es consumido debido a sus propiedades nutricionales, representando una gran opción, principalmente por su alto contenido proteico (12).
En el mundo, son producidas 14,2 millones de toneladas anualmente, en 14,8 millones de hectáreas, con una productividad de 0,96 t ha-1, (13), siendo la India el primer país en términos de producción y productividad.
Existen numerosos estudios acerca de la tolerancia a la salinidad en este cultivo; sin embargo, con respecto a los mecanismos involucrados en la tolerancia, aún no hay nada claro (14). La fase reproductiva del garbanzo (15), es más sensible a la salinidad, que la fase vegetativa y se ha demostrado que, por ser más sensible a este fenómeno durante el período de floración, le crea inestabilidad en su desarrollo y baja productividad (16). Aunque existen diversos criterios sobre la variabilidad de esta especie al estrés salino (14) aún no se cumplen todas las expectativas, por lo que se tendrá que continuar trabajando en la mejora genética para este carácter (15,17–20). Por tal razón esta investigación se realizó con el objetivo de determinar la tolerancia a la salinidad de cultivares cubanos de garbanzo, a través de indicadores del desarrollo y la acumulación de iones en los diferentes órganos de la planta.
MATERIALES Y METODOS
La investigación se llevó a cabo en el cuarto de luces del departamento de Bioquímica y Fisiología Vegetal perteneciente al INCA, en el período comprendido desde el 25 de mayo hasta el 15 de julio de 2015, con un fotoperíodo de 12 horas luz y 12 horas de oscuridad, y una temperatura que osciló entre 26 oC durante el día y 18 oC en la noche, según las exigencias del cultivo. Los cultivares evaluados fueron, del tipo Kabuli: Nac-29 (N-29), N-5HA, Nac-27 (N-27), Nac-24 (N-24), Nac-30 (N-30), Nac-6 (N-6), del tipo Gulabi: JP-94 y uno de origen extranjero: Blanco Sinaloa-92 (B. Sinaloa-92). Todos procedentes del banco de germoplasma del Instituto de Investigaciones Fundamentales de Agricultura Tropical (INIFAT).
Las semillas fueron previamente tratadas con una solución de TMTD a razón de 4 g kg-1 de semillas, durante diez minutos y luego se lavaron con agua destilada estéril. Posteriormente fueron pregerminadas en arena sílice estéril a temperatura ambiente y a los cuatro días de germinadas las plántulas fueron trasplantadas para recipientes plásticos que tenían una capacidad de 700 g de suelo.
Estos cultivares fueron sometidos a un tratamiento control y a 50 mM de NaCl. Se preparó una disolución de NaCl, según los tratamientos y se aplicó directamente a la planta, a razón de 50 mL por cada maceta, hasta el drenaje.
Se utilizó un diseño completamente aleatorizado, con 16 tratamientos que consistían en: cultivares de garbanzo en condiciones no salinas y cultivares de garbanzo sometidos a 50 mM de NaCl.
Se empleó un suelo Ferralítico Rojo Lixiviado, según última clasificación genética de los suelos (21) y un contenido de materia orgánica de origen animal, con una proporción de 2:1. El estrés fue impuesto a los 15 días de germinadas las plántulas.
Al final del experimento se procedió a realizar las mediciones de las variables correspondientes:
Para el procesamiento de los resultados, se realizó un análisis de varianza de clasificación simple, basado en un modelo lineal de efectos fijos y en los casos donde existieron diferencias significativas entre las medias se compararon mediante la prueba de Duncan (p≤0,05). Para la comparación entre tratamientos salinos y no salinos se empleó la distribución teórica de probabilidades t -Student para variables cuantitativas continuas, usando un tamaño de muestra n<30. Para todos estos análisis se utilizó el paquete estadístico profesional IBM.SPSS.Statistics versión 22. Para la construcción de las gráficas se utilizó el paquete estadístico SigmaPlot versión 11.0.
RESULTADOS Y DISCUSIÓN
Como una primera aproximación para estudiar los efectos del estrés salino y las respuestas a la tolerancia de plantas de garbanzo a la salinidad, se expuso en condiciones estresantes a diferentes cultivares cubanos de garbanzo.
El análisis realizado en la variable altura de la planta mostró la existencia altamente significativa entre las variedades ante la salinidad (50 mM). Tal resultado evidencia la respuesta diferenciada de los cultivares, lo cual tiene un significado práctico importante ya que permite seleccionar, inicialmente, variedades con tolerancia, conforme a la conductividad eléctrica de los suelos disponibles; y a mediano plazo, posibles progenitores para programas de mejora genética (22).
Actualmente el monitoreo de la respuesta de cultivares disponibles, ha sido una alternativa práctica y económicamente viable para elevar el coeficiente de utilización de los suelos salinos en diversas regiones del mundo (23).
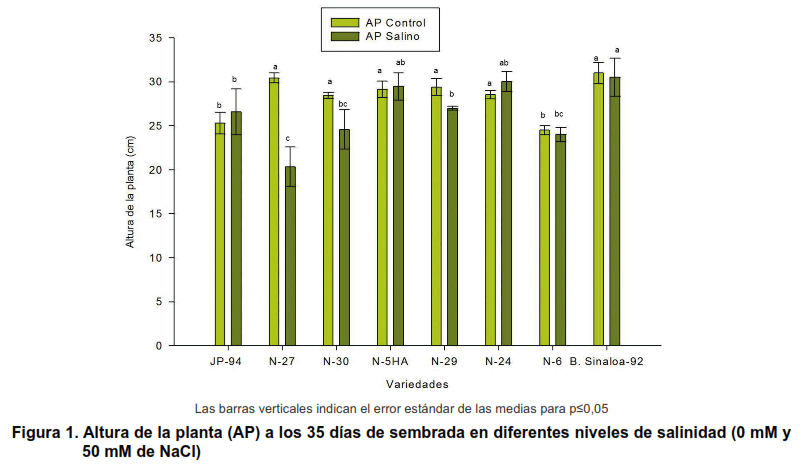
Al evaluar las diferencias en la altura de la planta en cada variedad (Figura 1) en las dos condiciones impuestas (0 y 50 mM de NaCl) se puede observar que este indicador es uno de los que más reducciones experimentaron durante el período de investigación.
En las condiciones experimentales de la presente investigación, ninguno de los cultivares cesó su crecimiento durante los primeros 15 días, luego de impuesto el estrés salino, casi todos los cultivares disminuyeron la tasa de crecimiento con respecto al control de cada variedad.
Los cultivares N-27, N-30, N-29, fueron las más impactadas por el efecto de la salinidad sobre este indicador; sin embargo, los cultivares N-5HA, N-24 y JP-94 fueron las que mejor respuesta manifestaron en condiciones salinas, cabe resaltar que exceptuando el cultivar Blanco Sinaloa-92, no manifestó reducciones significativas en esta variable, la variedad N-5HA, mostró un mejor crecimiento, con respecto al resto de los cultivares evaluados, ya que no tuvo reducción significativa del crecimiento.
En el presente ensayo, la reducción del crecimiento, quizás estuvo relacionada con alguna incapacidad que tienen las plantas de garbanzo para evitar que altas concentraciones de iones salinos lleguen a las hojas, aspecto que ha sido comprobado en diversas especies.
Los sistemas enzimáticos de la glicólisis son especialmente sensibles a las soluciones salinas, y dan como resultado una menor disponibilidad de energía, adquisición de nutrientes y una disminución del crecimiento de la planta (24).
A pesar de la sensibilidad del garbanzo a la salinidad, particularmente en los primeros estadios de desarrollo, existen algunos reportes de variabilidad interespecífica para este carácter. Los niveles más altos de concentraciones de sal en el suelo, conducen a una reducción significativa del desarrollo y a una pérdida de rendimiento de entre 8 y 10 %, a nivel mundial (25).
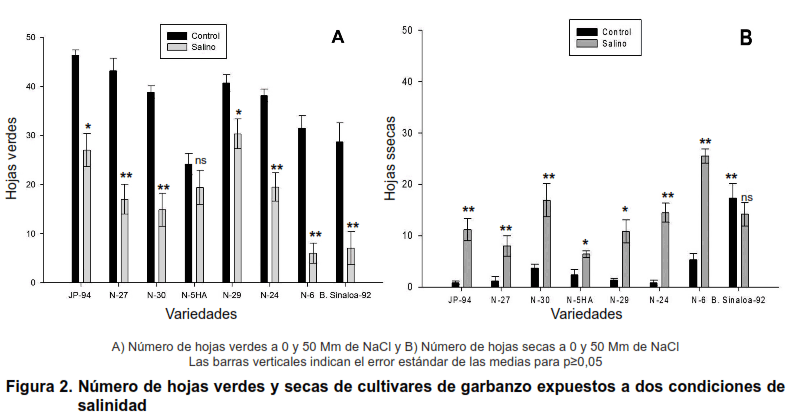
Otro indicador de gran importancia que se tuvo en cuenta para determinar la tolerancia a la salinidad, fue el número de hojas verdes y hojas secas, lo cual se evidencia en la Figura 2. Anteriormente se pudo observar que la variedad N-29 redujo un poco su crecimiento en cuanto a la altura, aunque no fue significativa esta reducción; sin embargo, en estos indicadores (número de hojas verdes y secas) fue la que mejor respuesta mostró, cuando fue comparada en el tratamiento salino, respecto al control.
En estos indicadores, los cultivares N-29, JP-94 y la N-5HA, fueron los menos afectados por sales, siendo este, un elemento que muestra, que estos cultivares podrían ser más tolerantes que el resto de los cultivares evaluados.
Por tanto, el número de hojas verdes se vio afectado por la salinidad, siendo más marcado el efecto en los cultivares Blanco Sinaloa-92, N-30 y N-6, a medida que aumentaba el tiempo de exposición a la salinidad.
En este estudio, los resultados evidencian que es posible que el Na+ se acumule, preferentemente, en las células epidérmicas de las hojas, y que esta sea expulsada al exterior de la hoja, reduciendo así la toxicidad por Na+.
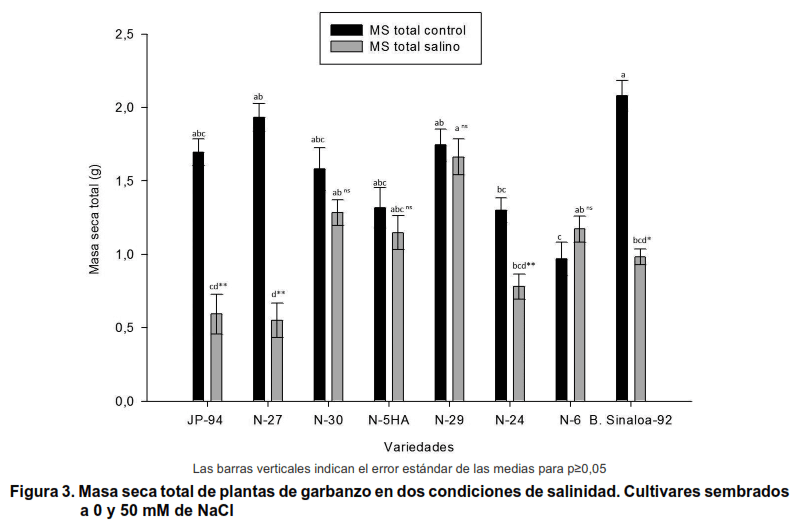
Con respecto a la masa seca total de la planta (Figura 3), se muestra que el cultivar N-29 manifiestómejor respuesta cuando fue expuesto a las condiciones salinas, se observó además, que mostró una mayor cantidad de biomasa en los tratamientos con sal y es el cultivar que menos se reduce ante estas condiciones, lo cual puede estar dado por una baja toxicidad del Na+ en los órganos de este cultivar.
Los cultivares en estudio manifestaron reducciones, con respecto a la biomasa de los diferentes órganos; sin embargo, en solo tres cultivares (N-29, N-5HA y N-30) se alcanzó un mayor contenido de masa seca en la raíz, tallo y hojas. Por otro lado, la figura muestra que el cultivar N-6 superó al resto de los cultivares, respecto a la masa seca total en condiciones salinas, tal resultado puede estar dado por el incremento en la producción de hojas de este cultivar, aunque se vio afectada negativamente por la salinidad, observándose un mayor número de hojas secas que verdes, pudiendo ser este uno de los cultivares más sensible al estrés salino en este estudio.
Al respecto, otros investigadores evaluando tolerancia a la salinidad de diferentes cultivares del tipo Kabuli y Desi, encontraron una reducción significativa de la biomasa cuando estas fueron expuestas a la salinidad, siendo esta mayor en el tratamiento control (20).
Por otro lado, otros utores, evaluando estrés salino hasta la floración y fructificación en diferentes líneas de garbanzo, encontraron una disminución en la biomasa de los rebrotes y la raíz (15).
En otros trabajos se evaluaron niveles de salinidad similar al presente ensayo en dos cultivares de garbanzo del tipo Desi, encontrándose reducciones en la masa seca de rebrotes y raíces con respecto al control y solo un cultivar se redujo al 6 % (26).
Las plantas pueden tolerar la salinidad mediante la exclusión de sodio y cloruro en las raíces, acumulando los iones en las hojas inferiores, de manera que la toxicidad de iones se evite en el crecimiento de hojas jóvenes y en el desarrollo de los tejidos reproductivos o tolerando los iones en los tejidos jóvenes y en desarrollo (4,19).
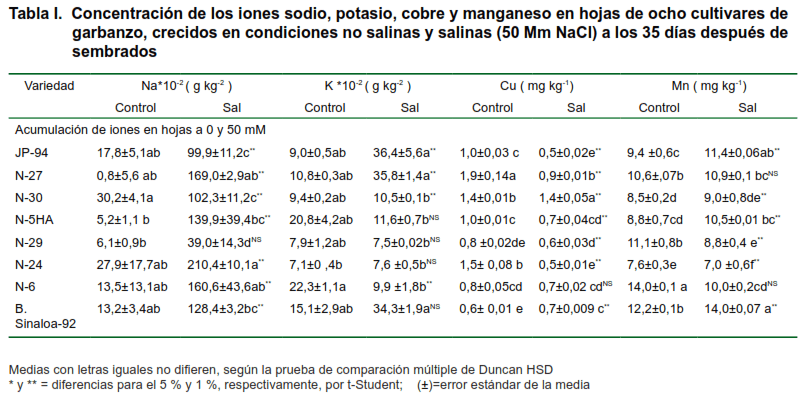
En la Tabla I se muestran las concentraciones de los iones sodio, potasio, cobre y manganeso en hojas de ocho cultivares cubanos de garbanzo. Se puede observar que los cultivares que resultaron ser tolerantes, teniendo en cuenta los indicadores de la biomasa, en algunas ocasiones acumularon menos cantidad de iones en los diferentes órganos que los otros genotipos sensibles.
En todos los cultivares evaluados, se incrementó la concentración de Na+, a pesar de que fueron muy variables en los diferentes órganos, siendo los cultivares N-27 y N-5HA los que mayores concentraciones de sodio acumularon en las hojas de este cultivo en las condiciones de salinidad impuesta.
Por otra parte, el cultivar N-29, resultó ser uno de los cultivares más tolerantes cuando se evaluaron las variables fisiológicas; al compararlo con el resto de los cultivares en ambas condiciones, se pudo observar que es uno de los que menos iones acumula en todos sus órganos, aunque su acumulación en las hojas, son inferiores, a las cantidades encontradas en el tallo y la raíz, para el tratamiento salino, esto pudo ser debido a que, al acumular la mayor cantidad de Na+ en los demás órganos (tallo y raíz), entraría menos cantidad de este ión tóxico al xilema y, como consecuencia, se acumularían menos cantidad en las hojas, pudiendo ser este un mecanismo fisiológico de tolerancia a la salinidad .
Sin embargo, el cultivar N-5HA que también muestra tolerancia en los demás indicadores, la acumulación de iones es variable en los diferentes órganos; por ejemplo, este cultivar donde mayor cantidad de Na+ acumula es en las hojas, mientras que en la raíz, cuando las comparamos con los otros cultivares, es uno de los que menos cantidad acumula, inferimos que estas variedades pueden presentar pelos excretores de sales y por el mecanismo de exclusión lleva estos iones a las hojas y los expulsa al exterior mediante estos pelos presentes en este órgano, ya que una alta toxicidad de este elemento reduce la fotosíntesis y reacciones enzimáticas.
Los demás iones (K, Cu y Mn) fueron menos acumulados por todos lo cultivares en las hojas, el cual está dado por las propias características de toxicidad que presenta el Na+.
Estos resultados coinciden con los obtenidos por otros investigadores (19,27), donde las concentraciones de Na+ en hojas de algunos genotipos fueron superiores a los de K+.
Se ha demostrado que el K+ es un elemento de gran importancia para este cultivo; sin embargo, se demostró que su acumulación disminuyó sustancialmente en los cultivares N-29 y N-5HA, alcanzándose los mayores valores en JP-94 y N-27. Cuando se analizaron las concentraciones de este ion en las hojas se pudo observar que el potasio se encontraba en mayor cantidad en el tratamiento salino para todos los cultivares en estudio, siendo estos valores inferiores en el tratamiento control.
Aunque se encontraba en menos cantidades que el Na+, había mayor acumulación de este ión en las hojas, cuando los comparábamos con el Cu y el Mn, ya que es un elemento que se traslada rápidamente desde las hojas viejas hasta las partes más nuevas.
La salinidad inducida disminuyó el contenido de estos iones en las hojas; sin embargo, el Mn se encontraba en mayor cantidad que el Cu y este se acumulaba más en la raíz que en los tallos y las hojas, siendo significativa su acumulación en el tratamiento salino que en el control. Además, debido a la poca movilidad el Cu, se pudo observar que este elemento se encontraba en pequeñas cantidades en las hojas, encontrándose una mayor disponibilidad de este elemento en las raíces.
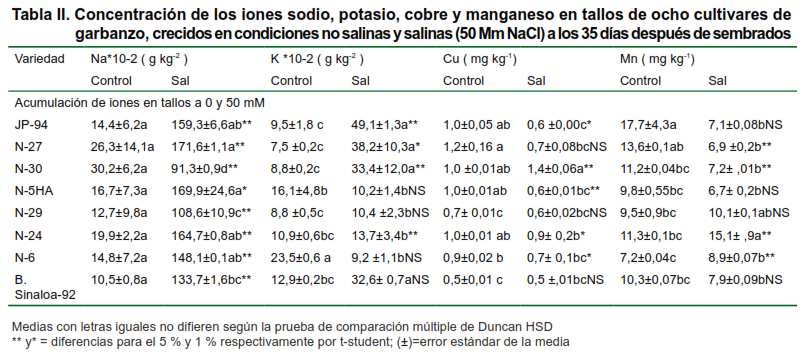
En la Tabla II se muestran las concentraciones de estos elementos anteriormente expuestos pero en el tallo, también en dos condiciones de salinidad.
Al analizar las concentraciones de sodio en este órgano para cada cultivar, se pudo observar que no hubo diferencia significativa en el tratamiento no salino; sin embargo, cuando estas plantas estuvieron sometidas a estrés por salinidad, se apreció una gran diferencia entre ellas, acumulándose menos cantidad de este elemento en las variedades N-29 y N-6. Por otro lado, el cultivar N-5HA, el cual ha venido mostrando cierta tolerancia, es uno de los cultivares que más acumula Na+ en el tallo cuando lo comparamos con el resto de los cultivares.
La entrada de K+ y Na+ en la célula se produce por la acción de transportadores y canales iónicos del plasmalema (28); sin embargo, existen transportadores muy selectivos para el K+, con una elevada afinidad por este elemento (10-50 mM), pero que también pueden transportar Na+ con baja afinidad y ser bloqueados por altas concentraciones de Na+ en medio (28).
Este fenómeno se puso de manifiesto en nuestra investigación, se puede observar en este órgano que el potasio aumenta en casi todos los cultivares en condiciones salinas; sin embargo, en los cultivares N-5HA y N-30, disminuye con respecto al control y estos mismos cultivares en la misma condición de salinidad impuesta, aumentan las concentraciones de Na+ en el tallo, por lo que coincidimos con los resultados encontrados (28).
El Cu y el Mn se encontraron en proporciones parecidas en el tallo de las diferentes variedades, siendo estos, cuantitativamente superiores en el tratamiento no salino.
Con respecto al Mn, se observó que en los cultivares N-29 y N-30 se acumula mayor cantidad de este elemento, cuando las plantas estaban sometidas a estrés por salinidad. De manera general, el Mn se encontraba más acumulado en el tallo que el Cu, debido a su translocación en este órgano, siendo, uno de los principales tejidos de reserva de este micronutriente, ya que al llegar a las hojas estos no pueden ser translocados. La baja acumulación de Cu puede estar dada también, a la poca movilidad que presenta este elemento en la planta.
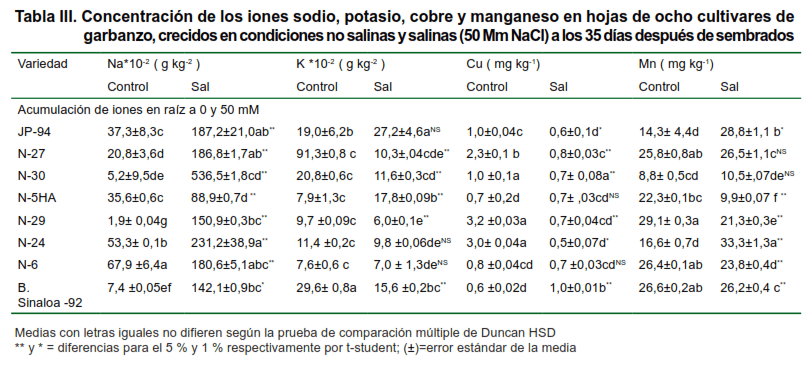
La raíz, como principal órgano de absorción de agua e iones, tiene gran importancia en la respuesta a corto y largo plazo al estrés salino.
Las características anatómicas y morfológicas de este órgano, pueden tener gran influencia en la capacidad de adaptación a la salinidad (28). En la Tabla III, se muestran los resultados encontrados de la acumulación de estos iones en este órgano.
Basado en el análisis iónico de la raíz, se puede observar que el resultado obtenido de los cuatro elementos en los diferentes cultivares estudiados, manifestaron respuestas diferentes cuando se comparaban los demás órganos de la planta.
Los niveles elevados de Na+, un ion principal en ambiente salino, puede inducir la deficiencia del elemento esencial K+, imponiendo un efecto antagónico sobre el K+, según investigaciones realizadas (29–31).
Estos resultados tuvieron igual tendencia a los obtenidos por estos autores (29–31), observándose una alta toxicidad del sodio en este órgano, trayendo como consecuencia disminución en la absorción del potasio por la raíz; aunque en el cultivar JP-94, en ambas condiciones, mostró menos acumulación de K+ que el Na+, este ión se encontraba en mayor cantidad en condiciones salinas que en el control.
En estudios recientes en este cultivo frente al estrés salino, se reportaron niveles bajos de Na+ en algunos órganos y tejidos reproductivos y se infiere que es poco probable que afecten adversamente los procesos reproductivos (15,32,33).
Los demás iones cobre y manganeso manifestaron una reducción significativa con respecto al sodio y al potasio y continúan estos cultivares mostrando mayor absorción de manganeso en condiciones de estrés, debido a que es la zona que más exporta este elemento donde se requiera y es donde primero se produce la afectación de su deficiencia. Los valores más bajos de acumulación de estos iones se observan en la variedad N-29 y N-5HA, con respecto al control, lo cual, puede estar dado por la alta toxicidad del sodio que hay presente en estos cultivares.
Estos resultados concuerdan con los obtenidos en otra investigación (33), donde se determinó la tolerancia a la salinidad en el cultivo del garbanzo.
CONCLUSIONES
La salinidad inhibió el desarrollo de los diferentes cultivares en estudio, los cultivares N-29, N-5HA del tipo Kabuli y el cultivar JP-94 del tipo Gulabi fueron los tolerantes, mientras que la N-6, N-24 y Blanco Sinaloa-92 los susceptible y dos cultivares moderadamente tolerantes N-27 y N-30.
La salinidad conllevó a variaciones significativas en las variables evaluadas, los cultivares tolerantes no mostraron diferencias en la altura de la planta, el número de hojas verdes, la biomasa de los diferentes órganos y la poca acumulación de iones en la raíz, el tallo y las hojas, cuando se comparaban con los cultivares susceptibles, donde el efecto acumulativo de estos cambios conducen a una mejor tolerancia al estrés.
BIBLIOGRAFÍA
1. Thakur P, Kumar S, Malik JA, Berger JD, Nayyar H. Cold stress effects on reproductive development in grain crops: An overview. Environmental and Experimental Botany. 2010;67(3):429–43. doi:10.1016/j.envexpbot.2009.09.004
2. Singh S, Parihar P, Singh R, Singh VP, Prasad SM. Heavy metal tolerance in plants: role of transcriptomics, proteomics, metabolomics, and ionomics. Frontiers in Plant Science. 2016;6(PMC4744854):1143. doi:10.3389/fpls.2015.01143
3. Roy SJ, Negrão S, Tester M. Salt resistant crop plants. Current Opinion in Biotechnology. 2014;26:115–24. doi:10.1016/j.copbio.2013.12.004
4. Zaki HEM, Yokoi S. A comparative in vitro study of salt tolerance in cultivated tomato and related wild species. Plant Biotechnology. 2016;33(5):361–72. doi:10.5511/plantbiotechnology.16.1006a
5. Martínez LA, Reyna IF, Payán JG, González EY, Aguilera JG. Efecto de la salinidad en callos de variedades de trigo durante el cultivo in vitro. Revista Mexicana de Ciencias Agrícolas. 2017;8(3):477–88.
6. Tavakkoli E, Fatehi F, Coventry S, Rengasamy P, McDonald GK. Additive effects of Na+ and Cl– ions on barley growth under salinity stress. Journal of Experimental Botany. 2011;62(6):2189–203. doi:10.1093/jxb/erq422
7. Álvarez A, Bañoz R, Otero L. Salinidad y uso de aguas salinas para la irrigación de cultivos y forrajes en Cuba. Ciencia y Tecnología Ganadera. 2008;2(1):1–12.
8. Ashraf MY, Ashraf M, Mahmood K, Akhter J, Hussain F, Arshad M. Phytoremediation of Saline Soils for Sustainable Agricultural Productivity. In: Ashraf M, Ozturk M, Ahmad MSA, editors. Plant Adaptation and Phytoremediation [Internet]. Dordrecht: Springer Netherlands; 2010 [cited 2018 Aug 28]. p. 335–55. doi:10.1007/978-90-481-9370-7_15
9. Ashraf MY, Awan AR, Mahmood K. Rehabilitation of saline ecosystems through cultivation of salt tolerant plants. Pakistan Journal of Botany. 2012;44(Special Issue):69–75.
10. Munns R. Genes and salt tolerance: bringing them together: Tansley review. New Phytologist. 2005;167(3):645–63. doi:10.1111/j.1469-8137.2005.01487.x
11. Hussain K, Nisar MF, Majeed A, Nawaz K, Bhatti KH, Afghan S, et al. What molecular mechanism is adapted by plants during salt stress tolerance? African Journal of Biotechnology. 2010;9(4):416–22.
12. Aguilar-Raymundo VG, Vélez-Ruiz JF. Propiedades nutricionales y funcionales del garbanzo (Cicer arietinum L.). Temas Selectos de Ingeniería de Alimentos. 2013;7(2):25–34.
13. FAOSTAT. FAO Statistics Division [Internet]. Crops, Production; 2014. Available from: http://Faostat.fao.org
14. Flowers TJ, Gaur PM, Gowda CLL, Krishnamurthy L, Samineni S, Siddique KHM, et al. Salt sensitivity in chickpea. Plant, Cell & Environment. 2010;33(4):490–509. doi:10.1111/j.1365-3040.2009.02051.x
15. Pushpavalli R, Quealy J, Colmer TD, Turner NC, Siddique KHM, Rao MV, et al. Salt stress delayed flowering and reduced reproductive success of chickpea (Cicer arietinum L.), a response associated with Na+ accumulation in leaves. Journal of Agronomy and Crop Science. 2016;202(2):125–38. doi:10.1111/jac.12128
16. Hussain N, Aslam M, Ghaffar A, Irshad M, Din N. Chickpea genotypes evaluation for morpho-yield traits under water stress conditions. JAPS: Journal of Animal & Plant Sciences. 2015;25(1):206–11.
Vadez V, Krishnamurthy L, Serraj R, Gaur PM, Upadhyaya HD, Hoisington DA, et al. Large variation in salinity tolerance in chickpea is explained by differences in sensitivity at the reproductive stage. Field Crops Research. 2007;104(1–3):123–9. doi:10.1016/j.fcr.2007.05.014
17. Krishnamurthy L, Turner NC, Gaur PM, Upadhyaya HD, Varshney RK, Siddique KHM, et al. Consistent variation across soil types in salinity resistance of a diverse range of chickpea (Cicer arietinum L.) genotypes. Journal of Agronomy and Crop Science. 2011;197(3):214–27. doi:10.1111/j.1439-037X.2010.00456.x
18. Turner NC, Colmer TD, Quealy J, Pushpavalli R, Krishnamurthy L, Kaur J, et al. Salinity tolerance and ion accumulation in chickpea (Cicer arietinum L.) subjected to salt stress. Plant and Soil. 2013;365(1–2):347–61. doi:10.1007/s11104-012-1387-0
19. Hirich A, El Omari H, Jacobsen S-E, Lamaddalena N, Hamdy A, Ragab R, et al. Chickpea (Cicer arietinum L.) physiological, chemical and growth responses to irrigation with saline water. Australian Journal of Crop Science. 2014;8(5):646–54.
20. Hernández A, Pérez J, Bosch D, Castro N. Clasificación de los suelos de Cuba. Mayabeque, Cuba: Ediciones INCA; 2015. 93 p.
21. Argentel Martínez L, Garatuza Payán J, González Y, A E, de los Santos- S. Evaluación de la tolerancia de variedades mexicanas de trigo a la salinidad, a través de indicadores fisiológicos, bioquímicos y agronómicos, cultivadas en Cuba en condiciones de campo. Cultivos Tropicales. 2016;37(1):91–101.
22. Blum A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production: Osmotic adjustment and plant production. Plant, Cell & Environment. 2017;40(1):4–10. doi:10.1111/pce.12800
23. Mata-Fernández I, Rodríguez-Gamiño M, López-Blanco J, Vela-Correa G. Dinámica de la salinidad en los suelos. Revista Digital del Departamento El Hombre y su Ambiente. 2014;1(5):26–35.
24. Arefian M, Vessal S, Bagheri A. Biochemical changes in response to salinity in chickpea (Cicer arietinum L.) during early stages of seedling growth. Journal of Animal and Plant Sciences. 2014;24(6):1849–57.
25. Khan HA, Siddique KHM, Colmer TD. Salt sensitivity in chickpea is determined by sodium toxicity. Planta. 2016;244(3):623–37. doi:10.1007/s00425-016-2533-3
26. Horie T, Karahara I, Katsuhara M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice. 2012;5(1):11. doi:10.1186/1939-8433-5-11
27. Leidi EO, Pardo JM. Tolerancia de los cultivos al estrés salino : qué hay de nuevo. Revista de Investigaciones de la Facultad de Ciencias Agrarias. 2002;2(2):70–91.
28. Kao CH. Mechanisms of salt tolerance in rice plants: Compatible solutes and aquaporins. Crop Environ. Bioinform. 2015;12:73–82.
29. Kao CH. Mechanisms of salt tolerance in rice plants: reactive oxygen species scavenging-systems. Journal Taiwan Agricultural Research. 2017;66(1):1–8. doi:10.6156/JTAR/2017.06601.01
30. Solangi SB, Chachar QI, Chachar SD, Solangi AB, Solangi JA. Effect of salinity (NaCl) stress on physiological characteristics of rice (Oryza sativa L.) at early seedling stage. International Journal of Agricultural Technology. 2016;12(2):263–79.
31. Kotula L, Khan HA, Quealy J, Turner NC, Vadez V, Siddique KHM, et al. Salt sensitivity in chickpea (Cicer arietinum L.): ions in reproductive tissues and yield components in contrasting genotypes: Growth and ions in reproductive organs of chickpea. Plant, Cell & Environment. 2015;38(8):1565–77. doi:10.1111/pce.12506
32. Atieno J, Li Y, Langridge P, Dowling K, Brien C, Berger B, et al. Exploring genetic variation for salinity tolerance in chickpea using image-based phenotyping. Scientific Reports [Internet]. 2017 [cited 2018 Aug 29];7(1). doi:10.1038/s41598-017-01211-7
Recibido: 06/12/2017
Aceptado: 16/06/2018
Yanitza Meriño-Hernández. Departamento de Producción Agrícola. Universidad de Granma, carretera a Manzanillo km 17.5, Bayamo, Granma. Código Postal 85 100. Email: yani@udg.co.cu
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}