Inoculación de hongos micorrizógenos arbusculares (HMA) y relaciones suelo pardo-abonos orgánicos en la aclimatización de vitroplantas de banano
Inoculation of arbuscular mycorhizal fungi (AMF) and Brown soil-organic fertilizers ratio in the acclimatization of banana vitroplants
Dr.C. Jaime E. Simó-González,I Dr.C. Luis A. Ruiz-Martínez,I Dr.C. Ramón Rivera-EspinosaII
IInstituto de Investigaciones de Viandas Tropicales (INIVIT), Departamento de Fitotecnia, Santo Domingo, Villa Clara, Cuba, CP 53000.
IIInstituto Nacional de Ciencias Agrícolas, gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700.
RESUMEN
A partir de la factibilidad y beneficios de la inoculación de cepas eficientes de hongos micorrízicos arbusculares en diferentes cultivos y el carácter micotrófico del banano, se realizó este trabajo con el objetivo de seleccionar cepas eficientes de HMA y establecer las relaciones suelo:abono orgánico (S:AO) óptimas para alcanzar una micorrización efectiva en la fase de aclimatización de plantas in vitro de bananos, cv. ‘FHIA-18’ (AAAB) en suelo Pardo mullido carbonatado. Se estudiaron cuatro cepas de HMA, dos tipos de abonos orgánicos (humus de lombriz y compost) y cinco relaciones S:AO: 100 % suelo, 3:1, 1:1, 1:3 y 100 % AO, utilizando un diseño Completamente Aleatorizado. El experimento se realizó en bolsas de 0,5 kg y se repitió dos veces y en cualquiera de las relaciones estudiadas los pH oscilaron entre 7,3 y 7,8. Se encontró una respuesta positiva y reproducible en los dos años a la inoculación de la especie Rhizoglomus intraradices (INCAM-11) en el crecimiento, estado nutricional, supervivencia y funcionamiento micorrízico de las plantas, con un comportamiento significativamente superior al resto de las cepas de HMA inoculadas. Asimismo, los efectos fueron dependientes de la relación S:AO, obteniéndose los mayores beneficios en presencia de las relaciones 1:1 y 1:3, cantidades inferiores de AO limitaron el funcionamiento micorrízico y superiores lo inhibieron. No se encontraron diferencias entre los tipos de AO estudiados. La utilización de la cepa R. intraradices (INCAM-11) en presencia de la relación 1:1 S:AO disminuyó en 50 % el contenido de AO de la recomendación vigente para la aclimatización de vitroplantas de banano.
Palabras clave: adaptación, compost, humus, micorrizas arbusculares vesiculares.
ABSTRACT
From the feasibility and benefits of inoculation of efficient strains of arbuscular mycorrhizal fungi in different cultures and mycotrophic character of bananas, this work was establish ratio soil: compost optimal ratios (S:AO, according its acronyms in English) for effective mycorrhization in the acclimatization phase of in vitro banana plants of banana cv. ‘FHIA-18’ (AAAB) in Fluffy brown carbonated soil. Four AMF strains, two sources of organic fertilizers (vermicompost and compost) and five S:AO ratio: 100 % soil, 3:1, 1:1, 1: 3 and 100 % AO were studied, using a Completely Randomized design. The experiment was performed in bags of 0,5 kg and it was repeated twice and in any of the ratio studied. The pH ranged between 7,3 and 7,8. A positive and reproducible response to Rhizoglomus intraradices (INCAM-11) inoculation were found after two years on growth, nutritional status, survival and mycorrhizal plant functioning with a significantly higher performance than other AMF inoculated strains. Also, the effects were dependent of the ratio S:AO, being obtained the biggest benefits in presence of the relationships 1:1 and 1:3, inferior quantities of AO limited the operation mycorrhizal and superiors inhibited it. No differences were found between AO sources studied. The use of R. intraradices (INCAM-11)straim in presence of the ratio 1:1 S:AO diminished in 50 % the content of AO of the effective recommendation for the acclimatization of banana vitroplants.
Key words: adaptation, composts, humus, vesicular arbuscular mycorrhizae.
INTRODUCCIÓN
La simbiosis micorrízica es una interacción biológica que se establece entre ciertos hongos del suelo y la mayoría de las especies vegetales (1). Los beneficios derivados de esta simbiosis se asocian con incrementos en la capacidad de absorción de agua y nutrimentos, mejoras en algunas propiedades del suelo, mayor tolerancia al déficit hídrico y disminución del daño de algunos patógenos foliares o radicales (2).
La inoculación con cepas eficientes de hongos micorrízicos arbusculares (HMA) en la producción agrícola, no es una práctica común internacionalmente (3,4), aunque en Cuba se dispone de una amplia información experimental para el manejo efectivo de cepas de HMA para un grupo extenso de cultivos como yuca
(Manihot esculenta Crantz), boniato (Ipomoea batata L.), tabaco (Nicotiana tabacum L.), especies de Brachiaria, maíz (Zea mays L.), frijol (Phaseolus vulgaris L.) entre otros y en diferentes ambientes edáficos (5-9), e incluso validados a escala productiva (10,11).
El programa de trabajo cubano se basa en un grupo de cepas generalistas con relación a las especies vegetales (12). Se ha encontrado que el manejo efectivo de la inoculación, depende del tipo de suelo o ambiente edáfico en que esta se efectúa (9), así como de aplicar una cantidad óptima de nutrimentos, vía fertilización mineral u orgánica, que permita que con la inoculación de la cepa eficiente de HMA para ese suelo se alcance una micorrización efectiva, siendo estas cantidades de fertilizantes menores que cuando no se inocula directamente el cultivo (13).
El banano y el plátano (Musa spp) son especies micotróficas y se han publicado algunos artículos sobre los efectos positivos de la inoculación con HMA en la fase de aclimatización de plantas de banano obtenidas in vitro (14).
En Cuba, en el caso del banano, se recomienda aclimatar las plantas obtenidas in vitro en sustratos conformados solo con abono orgánico (15), los cuales por la alta cantidad de nutrimentos que pueden liberar, no deben ser adecuados para un óptimo funcionamiento micorrízico (13).
Por todo lo anterior se realizó este trabajo con el objetivo de establecer la conducta y efectividad de la inoculación de cepas de HMA en la aclimatización de plantas de banano cv. ‘FHIA 18’ obtenidas in vitro, su dependencia con los tipos de AO y relaciones suelo:abonos orgánico en sustratos conformados con suelo Pardo mullido carbonatado, así como una comparación con la tecnología actual de aclimatización.
MATERIALES Y MÉTODOS
La investigación se llevó a cabo en las instalaciones de la Biofábrica de la Unión Agropecuaria Militar (UAM) ubicada en Santo Domingo, Villa Clara, Cuba durante los periodos mayo-julio de 2005 y mayo-julio de 2006 con plantas obtenidas in vitro de banano cv.’FHIA-18’ en la fase de aclimatización.
En el experimento se utilizaron suelos del tipo Pardo mullido, según la Clasificación de los suelos de Cuba 2015 (16), y catalogado asimismo como Phaeozems haplic calcaric en correspondencia con la World Reference Base (17). La caracterización del suelo se realizó de acuerdo a lo descrito por Paneque y colaboradores(18) (Tabla I).
Los suelos presentaron una reacción ligeramente alcalina, con valores de materia orgánica bajos y contenidos medios de fósforo disponible. En relación con los cationes intercambiables, el Ca mostró valores altos, mientras que el Mg y el K presentaron contenidos medios.
En cuanto al número inicial de esporas de HMA en el suelo (Tabla I), si bien fueron bajas, se corresponden con los valores obtenidos anteriormente por Ruiz y colaboradores (5) en este tipo de suelo y en esta misma localidad.
La aclimatización de las plantas de banano se realizó en condiciones semicontroladas, en una casa de cultivo cubierta por una malla sombreadora (marca Zarán), que permitía el paso del 60 % de la iluminación natural, con un flujo de fotones fotosintéticos (FFF) de 600 mol m-2 s-1 (20). Durante los 60 días de aclimatización se garantizó una temperatura entre 20 y 28 °C y una humedad relativa entre 80 y 90 % (19).
Se evaluaron cuatro cepas de HMA y un control sin inoculación, dos tipos de AO (compost y humus de lombriz) y cinco relaciones suelo-abono orgánico (v:v) [100 % Suelo (S), 3:1, 1:1, 1:3 y 100 % abono orgánico (AO)] en un diseño Completamente aleatorizado con 50 tratamientos, producto de todas las combinaciones de los diferentes factores evaluados. Para las evaluaciones se utilizaron 15 plantas por tratamiento. El tratamiento de 100 % AO con una u otra fuente correspondió al utilizado actualmente en la aclimatización de las plantas de banano obtenidas in vitro (15).
Las cepas utilizadas pertenecen a la colección del Instituto Nacional de Ciencias Agrícolas (INCA) de Cuba y se correspondieron con las siguientes especies: Funneliformis mosseae (20) / INCAM-2; Glomus cubense (21) / INCAM-4; Claroideoglomus claroideum (20) / INCAM-8 y Rhizoglomus intraradices (22) / INCAM-11. En todos los casos, el contenido de esporas de los inóculos se encontró entre 25 y 30 esporas g-1de producto.
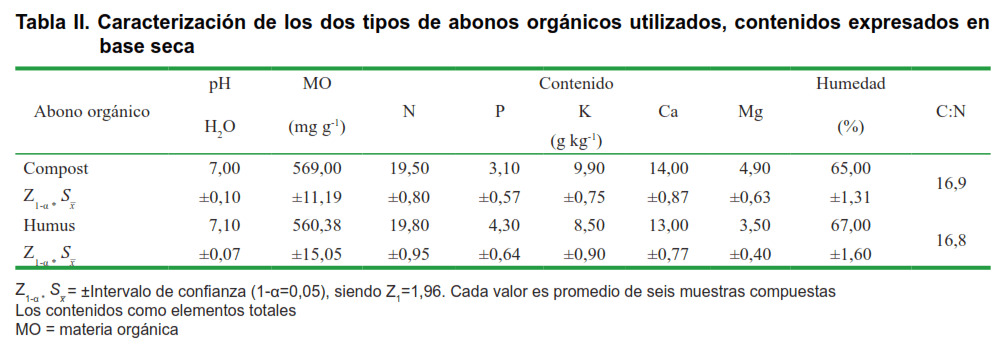
Los dos tipos de AO utilizados (compost y humus de lombriz) se caracterizaron como elementos totales (Tabla II) y las diferentes relaciones S:AO estudiadas como elementos disponibles o intercambiables
(Tabla III), según lo descrito por Paneque et. al. (18).
Para la conformación de las diferentes relaciones suelo:abono orgánico (S:AO), los tipos de AO se mezclaron independientemente con suelo Pardo mullido carbonatado (Tabla I) tomado a una profundidad de 0-20 cm.
Se utilizaron plantas de banano del cv. ‘FHIA-18’ (AAAB) obtenidas in vitro sin presencia de signos visibles de contaminación microbiana, con una altura aproximada de 50 mm y con 4 hojas por planta.
El trasplante se efectuó colocando una planta en cada bolsa de polietileno negro que ya contenía
0,5 kg de cada relación de S:AO (combinación de suelo y fuente de abono orgánico). La inoculación se realizó en el momento del trasplante a razón de 10 g de inóculo micorrízico por planta, localizado debajo de las raíces y en contacto directo con estas. El riego y las restantes actividades se desarrollaron acorde a lo establecido en el Instructivo Técnico (15).
El experimento tuvo una duración de 60 días y se evaluaron en ese momento, el porcentaje de supervivencia, la altura (cm), el área foliar (AF) (cm2), la masa seca (MS) (g), la concentración de N, P y K (g kg-1), la extracción de N, P y K (mg por planta) y el porcentaje de colonización micorrízica total.
Evaluaciones y determinaciones realizadas
Análisis inicial de esporas: para la extracción de esporas se procedió según el protocolo descrito por Gerdemann y Nicholson (23), modificado por Herrera y et. al. (24). Las esporas fueron contadas con el uso del microscopio estéreo (Carl Zeiss, Stemi 2000-C/50x) y se expresaron en esporas 50 g-1 de suelo.
Porcentajes de colonización micorrízica total: se determinó en cada muestra a partir de 200 mg de raíces, las cuales fueron secadas en estufa controlada termostáticamente, con ventilación forzada a 70 ºC hasta masa constante. Las raíces fueron teñidas según la metodología descrita por Rodríguez et. al. (25).
La evaluación se realizó en microscopio estereo (Carl Zeiss, Stemi 2000-C/50x). Para la cuantificación se utilizó el método de los interceptos de Giovanetti y Mosse (26).
Evaluaciones morfológicas: la altura (cm) se determinó con una regla graduada desde la base del seudotallo hasta la intercepción de las dos últimas hojas y el área foliar (AF, cm2) se calculó por la fórmula:
AF = largo x ancho x número de hojas x 0,80 x 0,662 (27).
Biomasa y concentración de (N, P y K): la masa fresca del tejido aéreo (MF) de cada planta se determinó por pesada en balanza técnica (± 0,01 g). La masa seca (MS) en cada muestra se determinó a partir del porcentaje de masa seca del tejido aéreo fresco secado en estufa controlada termostáticamente, con ventilación forzada a 70 ºC hasta masa constante y de los valores de masa fresca y se expresó en g.
En cada una de las muestras de tejido aéreo seco se determinaron las concentraciones totales de N, P y K (g kg-1), por el método descrito por Paneque et. al. (18). La extracción de N, P y K
(mg planta) se calculó a partir de las concentraciones de estos elementos en el tejido aéreo seco de la planta y la masa seca (g planta).
Análisis estadístico
Se verificaron en cada caso los supuestos de normalidad y homogeneidad de varianza por el paquete estadístico IBM SPSS (28), versión 11.5. En algunas de las variables evaluadas se establecieron los Intervalos de Confianza de las medias (1-α=0,05).
En cada uno de los dos años, con todas las variables y utilizando los 50 tratamientos se realizaron Análisis de Componentes Principales (ACP) y para establecer diferencias significativas entre los agrupamientos de tratamientos el Factorial Discriminante (FD) y el ANOVA a las puntuaciones discriminantes de las funciones. Una vez establecida la cepa eficiente se procedió a evaluar el efecto de los dos tipos de AO y las cinco relaciones S:AO en la efectividad de la inoculación micorrízica, procesando la información como un diseño completamente aleatorizado con arreglo factorial de 2x2x5x2 y solo utilizando la información asociada con la inoculación de la cepa eficiente y los homólogos no inoculados. Los factores y sus respectivos niveles en este caso fueron: cepas (2), tipo de AO (2), relaciones S:AO (5) y años (2).
En el caso de existir diferencias significativas, se utilizó como criterio de comparación entre medias la prueba de rangos múltiple de Tukey (p ≤ 0,05).
RESULTADOS
Selección de la cepa de hma más eficiente
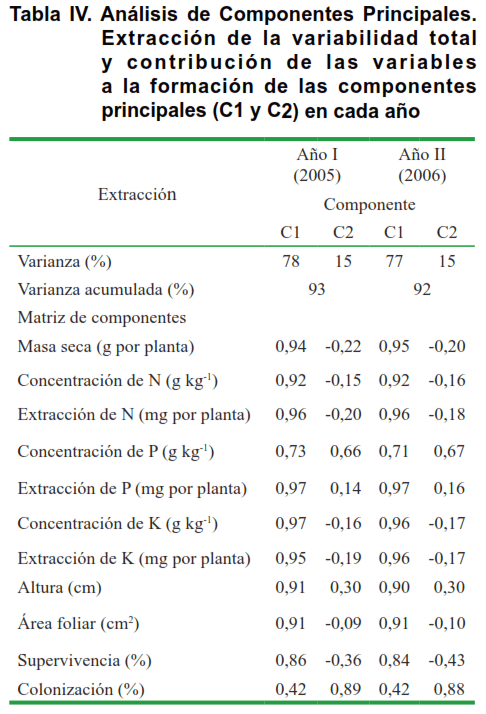
El ACP, mostró que los dos primeros componentes explicaron el 93 y 92 % de la varianza originada por los tratamientos, en cada uno de los años analizados (Tabla IV), por tanto resultó un método estadístico efectivo para explicar los efectos obtenidos por los tratamientos a partir de solo dos componentes.
El componente 1 presentó una participación alta y positiva de todas las variables, con excepción del porcentaje de colonización micorrízica que mostró los valores menores.
En el componente 2 con menor peso que el componente 1, solo el porcentaje de colonización micorrízica mostró una contribución alta e importante. La similar contribución de las variables evaluadas a la formación de los componentes en cada uno de los dos años, indicó una reproducibilidad alta de los resultados.
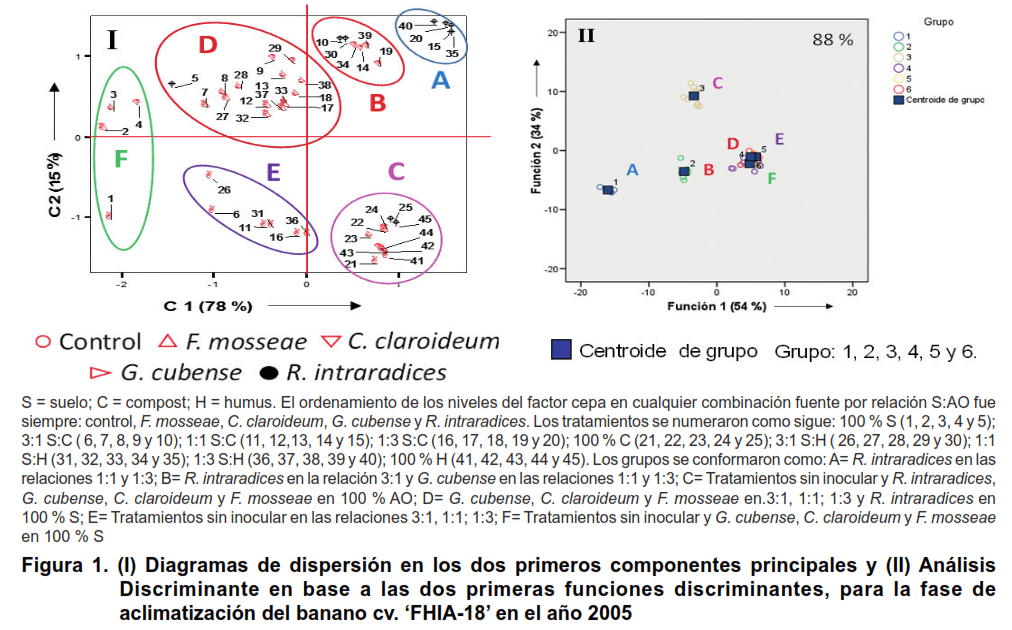
A partir de lo anterior, en las gráficas resultante del ACP se conformaron a priori seis grupos en cada uno de los dos años, los cuales se identificaron con las letras A, B, C, D, E y F y compuestos en ambos casos por los mismos tratamientos, expresión asimismo del carácter regular de los resultados. A partir de lo anterior se presentan los resultados del año 2005 (Figura 1 I).
Se destacó el grupo A, integrado por los tratamientos inoculados con la cepa de R. intraradices (INCAM-11) en presencia de las relaciones 1:1 y 1:3 de S:AO, bien fueran con una u otra fuente de abono orgánico, que presentaron siempre los valores máximos de ambos componentes, e indicando de acuerdo a la participación de las variables en los dos componentes (Tabla IV), que las plantas más vigorosas, con mayor tamaño y mayores contenidos nutricionales, mostraron también un funcionamiento micorrízico superior y una supervivencia mayor.
Cercano al grupo A, pero con un comportamiento inferior, y con valores positivos en ambos componentes, se encuentra el grupo B que incluyó los tratamientos inoculados con la cepa de R. intraradices en la relación 3:1 de S:AO y los inoculados con la cepa de G. cubense en las relaciones 1:1 y 1:3 de S:AO.
La inoculación con la cepa de G. cubense en estas relaciones de S:AO, mostró también un comportamiento beneficioso en las plantas de banano, aunque con valores inferiores en ambos componentes al que presentó R. intraradices en las relaciones 1:1 y 1:3 de S:AO, lo que indicó crecimiento, contenidos nutricionales y porcentajes de colonización micorrízica menores que los obtenidos con R. intraradices en esas combinaciones S:AO.
Por otra parte, se halla el grupo C, ubicado en el cuadrante inferior de la derecha con valores positivos del componente 1 y negativos del componente 2. Este grupo está compuesto por los tratamientos en la relación del 100 % AO (15) con cualquiera de los dos tipos de AO, incluyendo tanto los inoculados, con todas las cepas evaluadas, como los sin inocular. Este conjunto presentó los valores menores del componente 2, lo cual está asociado a un funcionamiento micorrízico bajo en estos tratamientos y a la vez presentó valores intermedios del componente 1.
El grupo D se ubicó en el cuadrante superior de la izquierda con valores negativos del componente 1
y positivos del componente 2. En el mismo se encuentran los tratamientos inoculados con las cepas de G. cubense, C. claroideum y F. mosseae en las relaciones 3:1, 1:1; 1:3 de S:AO, con una u otra fuente de abono orgánico y el inoculado con la cepa de R. intraradices en la relación del 100 % S.
Los tratamientos sin inocular en las relaciones 3:1, 1:1 y 1:3 de S: AO que conforman el grupo E, se ubicaron en el cuadrante inferior de la izquierda con valores negativos en ambos componentes.
Por último, se encuentra el grupo F que presentó los valores menores del componente 1 y asimismo valores bajos del componente 2. Este grupo lo conformaron los tratamientos que no recibieron abono orgánico (100 % S) incluyendo los inoculados y los no inoculados, con excepción del tratamiento inoculado con la cepa de R. intraradices que presentó siempre un comportamiento mejor en ambos componentes y se incluyó en el grupo D.
El AD permitió verificar la precisión del agrupamiento realizado a priori (Figura 1 II). Se encontró que en cada uno de los años con solo dos funciones se explicó el 88 y 90 % de las diferencias entre grupos, respectivamente. Teniendo en cuenta la alta reproducibilidad obtenida se presentan los resultados del AD del año 2005.
El ANOVA realizado a las puntuaciones discriminantes de la función 1 para los diferentes grupos, presentó diferencias significativas (p ≤ 0,01) entre estos, en los dos años (Tabla V), estableciendo el comportamiento siempre superior del grupo A conformado por los tratamientos que recibieron la inoculación con la cepa de R. intraradices (INCAM-11) en presencia de las relaciones 1:1 y 1:3 de S:AO y para ambos tipos de AO estudiados.
Del análisis conjunto de esta información se estableció que los tratamientos con mejores resultados estuvieron siempre asociados con la inoculación de la cepa de R. intraradices y por tanto se comportó como la cepa más eficiente para el banano en las condiciones estudiadas, aunque sus efectos fueron dependientes de la relación S:AO.
En el siguiente epígrafe se detalla para cada variable cómo influyeron las relaciones de S:AO en la efectividad de la simbiosis micorrízica arbuscular, pero solo en función de la inoculación con la cepa eficiente para estas condiciones, R. intraradices (INCAM-11).
Influencia de los tipos de abono orgánico y relaciones suelo:abono orgánico en la efectividad de la inoculación con la cepa eficiente de HMA
El análisis factorial para las diferentes variables, incluyendo los años como un factor, arrojó que este último factor no fue significativo y estuvo asociado con la alta reproducibilidad encontrada, por lo que se presentan los resultados de la interacción de máximo orden que fuera significativa, que en este caso fue la de tercer orden: factor cepas x factor tipos x factor relaciones.
Se encontró un efecto significativo (p ≤ 0,01) de la inoculación con la cepa de R. intraradices en el comportamiento de las diferentes variables estudiadas, de la cuales teniendo en cuenta las altas relaciones obtenidas entre las variables (tabla IV), se presentan aquí los obtenidos con cuatro de estas (Figura 2).
De forma tal que con su aplicación en presencia de las relaciones 1:1 y 1:3 de S:AO respectivamente, se presentaron siempre los valores mayores de cualquiera de las variables con diferencias significativas (p ≤ 0,01) con el resto de los tratamientos. El efecto significativo de la inoculación se observó también en el tratamiento que solo estaba compuesto por el suelo (100 % S), pero no se manifestó o fue poco importante en los tratamientos en que la relación S:AO solo estaba compuesta por el 100 % AO (Figuras 1 y 2).
Asimismo, los tratamientos inoculados en las relaciones 1:1 y 1:3 de S:AO originaron plantas más vigorosas y con contenidos de nutrimentos mayores que el tratamiento utilizado actualmente en condiciones de producción, 100 % AO.
En los tratamientos no inoculados se encontró una respuesta significativa a la aplicación creciente del abono orgánico, con diferencias significativas entre los valores obtenidos en las diferentes relaciones de S:AO, más evidentes en la masa seca, el área foliar, el porcentaje de supervivencia y la concentración de K.
En relación con el porcentaje de colonización micorrízica la inoculación con la cepa eficiente de HMA, aún en ausencia de abono orgánico (100 % S), originó valores relativamente altos de colonización micorrízica, aunque menores que los encontrados en las relaciones 1:1 y 1:3 de S:AO, dejando claro que esta variable si bien fue dependiente de la inoculación se necesita un suministro de nutrimentos para obtener los mayores porcentajes de colonización micorrízica.
En presencia del 100 % AO los porcentajes de colonización micorrízica fueron muy bajos y casi similares a los encontrados en el tratamiento de 100 % S no inoculado. Esta variable, si bien reflejó el efecto de la inoculación, presentó un comportamiento diferente al resto de las variables.
Los tipos de AO presentaron similares características, las cuales no presentaron entre sí diferencias en los contenidos de nutrimentos, materia orgánica y el pH (Tabla III). Las restantes variables estudiadas presentaron un comportamiento similar con una respuesta significativa a la inoculación, y siempre superior en las relaciones S:AO 1:1 y 1:3 y sin presentar diferencias con los tipos de AO.
DISCUSIÓN
Si bien la inoculación de cepas de HMA como vía para obtener una simbiosis micorrízica arbuscular efectiva presenta un reconocimiento creciente (2,4,13), existen diferentes factores que no permiten su introducción en los diferentes sistemas de producción.
Entre las diversas causas se encuentran la carencia de inoculantes efectivos que se apliquen en dosis bajas (3), la no comprensión del papel de las propiedades del suelo como factor determinante en la conducta de las cepas de HMA (9,13,29,30) y la necesidad de incrementar el conocimiento público sobre la importancia de las micorrizas en la sostenibilidad de los agroecosistemas (31).
Los resultados mostraron que la inoculación con la cepa de R. intraradices (INCAM-11) originó los efectos mayores en: el crecimiento, el estado nutricional, los porcentajes de colonización micorrízica total y de supervivencia de las plantas de banano durante la fase de aclimatización.
Con la aplicación de esta cepa en las relaciones de S:AO de 1:1 y 1:3 se obtuvieron porcentajes de colonización micorrízica total ligeramente superiores al 60 %, los cuales en muchos cultivos han sido indicativos de un funcionamiento micorrízico efectivo (13).
Asimismo, los resultados corroboran no solo la dependencia micorrízica del banano (14), sino también que la inoculación con la cepa eficiente de HMA en esta etapa del cultivo y en las relaciones de S:AO adecuadas para una micorrización óptima, mejora la tecnología de aclimatización de las plantas micropropagadas al obtenerse plantas más vigorosas y con porcentajes de supervivencia mayores que cuando estas crecen en presencia solamente de abono orgánico (100 % AO), que es la recomendación actual para la aclimatización de las plantas de banano (15).
Un funcionamiento micorrízico óptimo requiere de un suministro adecuado de nutrimentos, cantidades por debajo de este limitan la micorrización y por encima la inhiben (8,13) y si bien de forma general estas relaciones de efectividad y disponibilidad de nutrimentos han sido más comúnmente establecidas con fertilizantes minerales (13), en los trabajos realizados en cafetos se utilizaron los abonos orgánicos como fuentes de nutrimentos (13).
En aquellos experimentos, las relaciones de S:AO mejores para la micorrización del cafeto oscilaron entre 5:1 y 7:1, dependientes del tipo de suelo y posiblemente de la fertilidad química asociada. Suministros de nutrimentos inferiores, en forma de abono orgánico, no permitieron un funcionamiento micorrízico adecuado y cantidades superiores lo inhibieron.
En los experimentos con banano, los efectos mayores y beneficiosos de la inoculación se encontraron con cantidades superiores de abonos orgánicos, siendo presumiblemente una consecuencia de un ritmo mayor de crecimiento del banano, el hecho de que las relaciones de S:AO óptimas encontradas en el presente trabajo (1:1 y 1:3) contengan cantidades mayores de AO que las obtenidas en cafeto.
Es decir, la influencia de la disponibilidad de nutrimentos en la efectividad de una cepa eficiente y en su funcionamiento, no es independiente del cultivo micorrizado y de sus requerimientos. Como la relación 1:1 de S:AO además requirió de una cantidad menor de AO, resultó más económica y por tanto a partir de estos resultados es la que se recomienda.
Los tratamientos inoculados o no, que dependieron solo de la fertilidad del suelo (100 % S) y de relaciones con cantidades menores de abono orgánico (3:1 de S:AO), no garantizaron un suministro adecuado de nutrimentos, lo que conllevó a porcentajes de colonización micorrízica inferiores, concentraciones nutricionales menores y crecimiento inadecuado en las plantas de banano.
Asimismo, la disminución del efecto de la inoculación en presencia de los tratamientos que solo recibieron abono orgánico (100 % AO) parece estar relacionada con el efecto previamente informado de inhibición de la micorrización en presencia de una disponibilidad alta de nutrimentos (8,13).
Es de destacar que las aplicaciones crecientes de abono orgánico no cambiaron la conducta respectiva de las cepas de HMA y de esta forma R. intraradices presentó un comportamiento superior a las restantes cepas en cualquier relación de S:AO, con excepción de la alcanzada en los tratamientos con 100 % AO en que prácticamente no existió efecto de ninguna de las cepas de HMA.
Lo anterior puede ser una consecuencia de las características de los abonos orgánicos utilizados y del propio suelo y de que en las diferentes relaciones de S:AO estudiadas, si bien estas difirieron en los contenidos de nutrimentos, mantuvieron valores de pH relativamente similares y superiores a siete (Tabla III).
Diversos autores señalaron la importancia del pH en el funcionamiento de la simbiosis micorrízica y la distribución de las cepas de HMA en los agroecosistemas (32,33). En un trabajo resumen de 35 experimentos en que se compararon la conducta de las mismas cepas de HMA que aquí se estudiaron, pero en diversos cultivos y suelos (9), se obtuvieron relaciones altas (R2=0,97) entre la efectividad de las cepas de HMA inoculadas y el pH del suelo. Estos autores informaron que el pH parece ser una de las propiedades fundamentales del suelo que determinan el cambio de efectividad de las cepas inoculadas de HMA al pasar de un suelo a otro.
Lo anterior explicaría por qué en las diferentes relaciones de S:AO, R. intraradices (INCAM-11) presentó efectos mayores que las restantes cepas y por tanto los resultados obtenidos en estas condiciones fueron similares a los logrados en diferentes experimentos y cultivos en suelos Pardos mullidos carbonatados (pH≥7), en los que siempre R. intraradices (INCAM-11) se comporta como la cepa de HMA más eficiente (9), con resultados superiores significativamente a los obtenidos cuando se inocularon otras cepas de HMA.
En el caso específico del efecto de las micorrizas arbusculares en presencia de abonos orgánicos y residuos vegetales, actualmente se reconoce la intensificación de la mineralización de los residuos orgánicos por la presencia de las micorrizas en el sistema suelo-planta y una absorción preferencial por la red de hifas micorrízicas (2), asociándose además con la proliferación de las hifas en los residuos y la interacción positiva con otros microorganismos del suelo que descomponen los restos orgánicos (34). Todos estos mecanismos de una u otra forma favorecen el aumento de la capacidad de absorción y disponibilidad de los nutrimentos, originalmente en formas orgánicas.
Si bien se encontró una respuesta creciente a la aplicación de dosis superiores de AO en ausencia de la inoculación, coincidente con la recomendación del Instructivo (15), los resultados de este trabajo demostraron que la inoculación con la cepa eficiente de HMA en la relación de S:AO de 1:1 garantizó no solo un funcionamiento micorrízico óptimo, sino también plantas más vigorosas, con mejores índices de crecimiento, estado nutricional y supervivencia y por tanto mejoró la tecnología vigente de aclimatización, disminuyendo además las cantidades de abonos orgánicos requeridas.
CONCLUSIONES
Se encontró una respuesta beneficiosa de las plantas de banano obtenidas in vitro a la inoculación con cepas de HMA en la etapa de aclimatización en sustratos a base de compost o humus y suelo Pardo mullido carbonatado, con una conducta diferenciada entre las cepas y con los mayores efectos obtenidos, tanto sobre el crecimiento, porcentajes de supervivencia, estado nutricional (g kg-1 de N, P y K) y porcentajes de colonización micorrízica con la aplicación de R. intraradices. Los mayores efectos se obtuvieron en presencia de las relaciones de 1:1 y 1:3 partes de suelo:abono orgánico. Cantidades menores limitaron la efectividad de la micorrización y superiores como 100 % AO la inhibieron.
No se encontraron diferencias entre los tipos de abono orgánicos evaluados. La cepa con mejor desempeño R. intraradices (INCAM-11) fue la misma que ha sido seleccionada para muy diversos cultivos en condiciones de suelos con pH≥7,0, similar condición a la encontrada en cualquiera de las relaciones suelo:abono orgánico estudiadas.
La utilización de la cepa R. intraradices (INCAM-11) en presencia de la relación 1:1 S:AO disminuyó en 50 % el contenido de AO de la recomendación vigente para la aclimatización de vitroplantas de banano.
BIBLIOGRAFíA
1. Smith SE, Smith FA. Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth: New Paradigms from Cellular to Ecosystem Scales. Annual Review Plant Biology. 2011;(62):227-50. doi: 10.1146/annurev-arplant-042110.
2. Priyadharsini P, Muthukumar T. Insight into the Role of Arbuscular Mycorrhizal Fungi in Sustainable Agriculture. In: Thangavel P, Sridevi G, editors. Environmental Sustainability. Springer India; 2015. doi: 10.1007/978-81-322-2056-5_1.
3. Verbruggen E, Heijden MGA, Rillig MC, Kiers ET. Mycorrhizal fungal establishment in agricultural soils: factors determining inoculation success. New Phytologist. 2012;197(4):1104–9. doi: 10.1111/j.1469-8137.2012.04348.x.
4. Yang WC, Ellouze A, Navarro-Borrell A, Esmaeili TR, Klabi M, Dai ZK, Hamel C. Management of the Arbuscular Mycorrhizal Symbiosis. Sustainable Crop Production. In: Solaiman ZM, editor. Mycorrhizal Fungi: Use in Sustainable Agriculture and Land Restoration. Heidelberg, Berlin: Springer-Verlag; 2014. p. 89-118. doi: 10.1007/978-3-662-45370-4-7.
5. Ruiz L, Simó J, Rodríguez S, Rivera R. Las micorrizas en cultivos tropicales. Una contribución a la sostenibilidad agroalimentaria. Editorial Académica Española 2012 239 p. ISBN: 978-3-8484-5382-5.
6. Cruz HY, García RM, Hernández MJM, León GY. Influencia de las micorrizas arbusculares en combinación con diferentes dosis de fertilizante mineral en algunas características morfológicas de las plántulas de tabaco. Cultivos Tropicales. 2012;33(3):23-6.
7. Martín G, Rivera R, Pérez A. Efecto de canavalia inoculación micorrízica y dosis de fertilizante nitrogenado en el cultivo del maíz. Cultivos Tropicales. 2013;34(4):60-7.
8. González CPJ, Ramírez PJF, Morgan RO, Rivera ER, Plana LR. Contribución de la inoculación micorrízica arbuscular a la reducción de la fertilización fosfórica en Brachiaria decumbens. Cultivos Tropicales. 2015;36(1):135-42.
9. Rivera R, González PJ, Hernández A, Martín G, Ruiz L, Fernández K, Simó J, García M, Pérez A, Riera M, Bustamante C, Joao JP, Ruiz M. La importancia del ambiente edáfico y del pH sobre la efectividad y la recomendación de cepas eficientes de HMA para la inoculación delos cultivos. In: VIII Congreso de la Sociedad Cubana de la Ciencia del Suelo. La Habana, Cuba; 2015.
10. Rivera R, Ruíz L, Riera M, Simó J, Fundora LR, Calderón A, Martín JV, Marrero Y, Joao JP. La efectividad del biofertilizante EcoMic® en el cultivo de la yuca. Resultados de las campañas de extensiones con productores. Cultivos Tropicales. 2012;33(1):5-10.
11. Rivera R, Calderón A, Nápoles MC, Ruiz L. La validación a escala productiva del biofertilizante EcoMic® y su aplicación conjunta con rizobios en el cultivo del frijol en el centro y occidente del país. Mayabeque, Cuba: INCA; 2012. 19 p. doi: 10.13140/2.1.4028.8320.
12. Opik M, Moura M. Missing nodes and links in mycorrhizal networks. New Phytologist. 2012;(194):304–6.
13. Rivera R, Fernández F, Fernández K, Ruiz L, Sánchez C, Riera M. Advances in the management of effective arbuscular mycorrhizal symbiosis in tropical ecosystesm. In: Hamel C, Plenchette C, editors. Mycorrhizae in Crop Production. Binghamton, N.Y.: Haworth Press; 2007. p. 151-96. doi: 10.13140/RG.2.1.1771.2162.
14. Jaizme-Vega MC, Delamo M, Tenoury P, Rodríguez-Romero AS. Efectos de la micorrización sobre el desarrollo de dos cultivares de platanera micropropagada. InfoMusa. 2002;11(1):25-8.
15. MINAG (Ministerio de la Agricultura). Instructivo Técnico para la producción de Viandas. Producción de semillas por método biotecnológico. 1st ed. La Habana, Cuba: Instituto de Investigaciones de Viandas Tropicales (INIVIT); 2012. p. 121-2.
16. Hernández A, Pérez JM, Bosch D, Castro N. Clasificación de los suelos de Cuba 2015. Mayabeque, Cuba: Ediciones INCA; 2015. 93 p.
17. IUSS Working Group WRB. World Reference Base for soil resources 2014: international soil classification system for naming soils and creating legends for soil maps. Rome: Food and Agriculture Organization of the United Nations; 2014. 191 p. (World Soil Reports).
18. Paneque PVM, Calaña NJM, Calderón VM, Borges BY, Hernández GTC, Caruncho CM. Manual de técnicas analíticas para análisis de suelo, foliar, abonos orgánicos y fertilizantes químicos [Internet]. 1st ed. La Habana, Cuba: Ediciones INCA; 2010 [cited 2016 Jan 27]. 157 p. Available from: http://mst.ama.cu/578/
19. Robinson JC, Galán SV. Plátanos y Bananas. España: Mundiprensa; 2012. 321 p.
20. Schüβler A, Walker C. Evolution of the ‘Plant-Symbiotic’ Fungal Phyllum Glomeromycota. Evolution of fungi and fungal-like organisms. In: Pöggeler S, Wöstemeyer J, editors. The Mycota XIV. Heidelberg, Berlin: Springer-Verlag; 2011. p. 163-85.
21. Rodríguez Y, Dalpé Y, Séguin S, Fernández K, Rivera R. Glomus cubense sp. nov. an arbuscular mycorrhizal fungus from Cuba. Micotaxon. 2011;118(1):337-47.
22. Sieverding E, Alves da Silva G, Berndt R, Oehl F. Rhizoglomus a new genus of the Glomeraceae. Mycotaxon. 2014;129(2):373 -86.
23. Gerdemann JW, Nicholson TH. Spore of mycorrhizae endogone species extracted from soil by wet sieving and decanting. Transactions of the British Mycological Society. 1963;46(2):235-44. doi: 10.1016/S0007-1536(63)80079-0.
24. Herrera RA, Ferrer RL, Furrazola E, Orozco MO. Estrategia de funcionamiento de las micorrizas VA en un bosque tropical. Mérida, México: Programa Iberoamericano de Ciencia y Tecnología para el desarrollo. Subprograma XII, Diversidad Biológica; 1995.
25. Rodríguez YY, Arias PL, Medina CA, Mujica PY, Medina GLR, Fernández SK, Mena EA. Alternativa de la técnica de tinción para determinar la colonización micorrízica. Cultivos Tropicales. 2015;36(2):18-21.
26. Giovanetti M, Mosse B. An Evaluation of Techniques for MeasuringVesicular Arbuscular Mycorrhizal Infection in Roots. New Phytologist. 1980;84(3):489-500. doi: 10.1111/j.1469-8137.1980.tb04556.x.
27. Kumar N, Krishnamoorthy V. Nuevo factor para determinar el área foliar en los bananos. InfoMusa. 2002;11(2):42-3.
28. IBM Corporation. IBM SPSS Statistics [Internet]. Version 11.5. U.S: IBM Corporation; 2012. Available from: http://www.ibm.com
29. Santos-González JCS, Nallanchakravarthula SA, Finlay RD. Soil but not cultivar shapes the structure of arbuscular mycorrhizal fungal assemblages associated with strawberry. Microbiology. Ecology. 2012;(62):25–35. doi: 10.1007/s00248-011-9834-7.
30. Sikes BA, Maherali H, Klironomos JN. Mycorrhizal fungal growth responds to soil characteristics but not host plant identity during a primary lacustrine dune succession. Mycorrhiza. 2014;24:219-26.
31. Gianinazzi S, Vosatka M. Inoculum of arbuscular mycorrihizal fungi for production systems: science metes business. Canadian Journal of Botany. 2004;82(8):1264-71.
32. Siqueira JO, Franco AA. Biotecnología do solo. Fundamentos e Perspectivas. Brasilia: MEC - ABEAS - Lavras - ESAL – FAEPE; 1988. 236 p.
33. Göransson P, Olsson PA, Postma J, Falkengren-Grerup U. Colonisation by arbuscular mycorrhizal and fine endophytic fungi in four woodland grasses-variation in relation to pH and aluminium. Soil Biology Biochemistry. 2008;40:2260-5. doi: 10.1016/j.soilbio.2008.05.002.
34. Hodge A, Storer K. Arbuscular mycorrhiza and nitrogen: implications for individual plants through to ecosystems. Plant and Soil. 2015;386:1-19. doi: 10.1007/s11104-014-2162-1.
Recibido: 13/04/2016
Aceptado: 13/02/2017
Dr.C. Jaime E. Simó-González, Instituto de Investigaciones de Viandas Tropicales (INIVIT), Departamento de Fitotecnia, Santo Domingo, Villa Clara, Cuba, CP 53000. Email: micorrizasf@inivit.cu