Contribución
de la inoculación micorrízica arbuscular a la reducción de la fertilización
fosfórica en Brachiaria decumbens
Contribution
of arbuscular mycorrhizal inoculation to the reduction of phosphorus fertilization
in Brachiaria decumbens
Ms.C.
Pedro J. González Cañizares,I Ms.C. Juan F. Ramírez Pedroso,II
Ms.C. Osvaldo Morgan Rosemond,II Dr.C. Ramón Rivera Espinosa,I
Dr.C. Rodolfo Plana LlerenaI
IInstituto
Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas,
Mayabeque. Cuba. CP 32 700.
IIInstituto de Investigaciones de Pastos y Forrajes, avenida Independencia,
km 8½ Boyeros, La Habana, Cuba.
RESUMEN
Se realizó un
experimento en un suelo Ferralítico Rojo Lixiviado, con el objetivo de evaluar
la contribución de la inoculación micorrízica arbuscular a la reducción de la
fertilización fosfórica en Brachiaria decumbens cv. Basilisk. Se estudiaron
dosis de 0, 30, 60 y 90 kg de P2O5 ha-1 año-1,
solas y combinadas con la inoculación de la cepa de hongo micorrízico arbuscular
Glomus cubense, seleccionada previamente por su alta eficiencia, en un diseño
de bloques al azar con arreglo factorial 4x2 y cuatro réplicas. El experimento
se condujo bajo condiciones de riego y tuvo una duración de dos años. La inoculación
de G. cubense incrementó significativamente los porcentajes de colonización
micorrízica, la densidad visual y el número de esporas en la rizosfera, aunque
los valores de estas variables disminuyeron con la dosis más alta de P2O5.
En los tratamientos no inoculados, los mayores rendimientos y extracciones de
P en biomasa aérea del pasto se obtuvieron con las dosis de 30 a 90 kg de P2O5
ha-1, sin diferencias significativas entre ellas; sin embargo, solo
con la inoculación de G. cubense, durante el primer año del establecimiento
del pasto ambos indicadores alcanzaron valores similares a los obtenidos con
las aplicaciones de fertilizante fosfórico sin inocular. La influencia de G.
cubense se observó hasta el período lluvioso del segundo año, aunque los valores
de las variables fúngicas, los rendimientos y las exportaciones de P en la biomasa
de la parte aérea indicaron una disminución de la efectividad de la inoculación,
en relación con el primero.
Palabras
clave: Glomus, nutrición, fertilización, gramíneas forrajeras.
ABSTRACT
An experiment
was conducted on a Red Ferralitic Lixiviated soil in order to assess the contribution
of arbuscular mycorrhizal inoculation to reduce phosphorus fertilization in
Brachiaria decumbens cv. Basilisk. Doses of 0, 30, 60 and 90 kg P2O5
ha-1 year-1, alone and combined with the inoculation of
arbuscular mycorrhizal fungal strain Glomus cubense, previously selected for
its high efficiency, were distributed in a randomized block design with 4x2
factorial arrangement and four replications. The experiment was conducted under
irrigated conditions and lasted two years. G. cubense significantly increased
mycorrhizal colonization percentages, visual density and spore number in the
rhizosphere, although the values of those variables decreased with the highest
doses of P2O5. In non inoculated treatments, the highest
yields and P extractions in the aerial biomass were obtained with doses of 30-90
kg of P2O5 ha-1, with no significant differences
between them, but only with the inoculation of G. cubense, during the first
year of the establishment of grass both indicators reached values similar to
those obtained with phosphoric fertilizer applications without inoculation.
The influence of G. cubense was observed until the rainy season of the second
year, although fungal variable values, yields and P exports with the aerial
biomass indicated a decrease in the effectiveness of inoculation, in relation
to the first year.
Key words:
Glomus, nutrition, fertilization, forage grasses.
INTRODUCCIÓN
La fertilización
de los pastos mejorados constituye una herramienta para incrementar la oferta
de forraje por unidad de superficie y tiempo y, consecuentemente, la producción
animal. Por esta vía, no solo se restituyen los nutrientes que se extraen del
suelo con la biomasa que consume el ganado, sino también se mejora el valor
nutritivo y la persistencia de los pastos (1).
Hasta hace pocos años, los estudios sobre la fertilización de los pastos se
enfocaban, principalmente, en el incremento de la producción de biomasa por
unidad de superficie y la evaluación del impacto económico del uso de los fertilizantes.
Actualmente, sin perder de vista el aspecto económico dado el aumento creciente
de los precios de los fertilizantes, se le concede gran importancia a la preservación
del ambiente; de modo que se ha acrecentado la necesidad de diseñar estrategias
de fertilización que garanticen una nutrición adecuada de los pastos y a la
vez, aseguren la protección de los recursos naturales (2, 3).
Dentro de esas estrategias se puede considerar el manejo de la simbiosis micorrízica,
ya que los beneficios de los hongos micorrízicos arbusculares (HMA) en los agroecosistemas
de pastizales están estrechamente relacionados con el aumento de la absorción
de elementos minerales, agua y otras sustancias, a través de una red de hifas
interconectadas que incrementan el volumen de suelo que exploran las raíces
y facilitan el acceso de las plantas a los nutrientes que se encuentran en formas
menos asimilables (4, 5). Así, podrían disminuirse las dosis de fertilizantes
a aplicar a los pastos en función de un mayor aprovechamiento y eficiencia del
uso de los nutrientes del suelo y de los propios fertilizantes.
Entre los nutrientes cuya absorción se favorece por la acción de los HMA, el
P es uno de los más importantes, pues entre un 95 y un 99 % de las cantidades
de este elemento presentes en el suelo se encuentran en formas no disponibles
para las plantas (6). En el caso específico de los agroecosistemas ganaderos
de las regiones tropicales, donde el bajo aprovechamiento del P por muchas especies
de gramíneas forrajeras está indisolublemente ligado a la compleja dinámica
de este elemento en los suelos (7, 8), el manejo de las asociaciones micorrízicas
puede constituir una opción válida para mejorar la eficiencia de la fertilización
fosfórica.
Basado en estos aspectos se realizó este trabajo, cuyo objetivo fue evaluar
la contribución de la inoculación de hongos micorrízicos arbusculares (HMA)
a la reducción de la fertilización fosfórica en Brachiaria decumbens cv. Basilisk.
MATERIALES
Y MÉTODOS
El experimento
se realizó en condiciones de campo, en la Microestación de Pastos y Forrajes
de la Empresa Pecuaria Genética “Niña Bonita”, ubicada en el municipio de Bauta,
provincia de Artemisa, sobre un suelo Ferralítico Rojo Lixiviado (9) cuyas principales
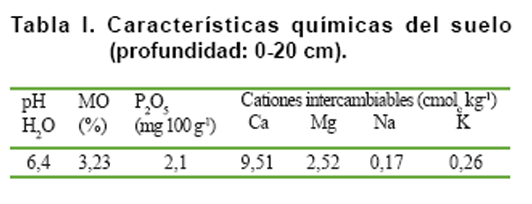
características químicas (Tabla
I), reflejaron un pH ligeramente ácido, un contenido medio de materia orgánica
y bajos tenores de P2O5 asimilable y K intercambiable
(10).
Para los
análisis del suelo se utilizaron los métodos establecidos en el laboratorio
de suelos y tejido vegetal del Departamento de Biofertilizantes y Nutrición
de las Plantas del Instituto Nacional de Ciencias Agrícolas (INCA) (11).
En un diseño experimental de bloques al azar con arreglo factorial 4x2 y cuatro
réplicas, se evaluaron cuatro dosis de fertilizante fosfórico (0, 30, 60 y 90
kg de P2O5 ha año-1) en dos condiciones de
inoculación micorrízica (sin inocular e inoculado con la cepa de HMA Glomus
cubense). Los tratamientos se distribuyeron en parcelas, que constituyeron la
unidad experimental, que tenían una superficie total de 28 m2 y un
área de cálculo de 21 m2.
El suelo se preparó mediante labores de roturación (arado), grada, cruce (arado)
y grada, a intervalos aproximados de 20 días entre cada una. La siembra del
pasto se realizó el 15 de marzo de 2005, en surcos separados a 70 cm y a chorrillo,
con una dosis de 6 kg de semilla total ha-1 (1 kg de semilla pura
germinable ha-1) y a una profundidad de 1,5 cm. El fertilizante fosfórico
se aplicó en las dosis indicadas para cada tratamiento, al momento de la siembra
y a los 12 meses posteriores. A todos los tratamientos se les aplicó un fondo
fijo de 210 kg de N ha-1 año-1, fraccionado en dosis de
35 kg ha-1 al momento de la siembra y después de cada corte y 100
kg de K2O ha-1 año-1, en una dosis única distribuida también
en la siembra y a los 12 meses. Como portadores se utilizaron el superfosfato
triple, la urea y el cloruro de potasio.
Para la selección de las dosis y frecuencias de aplicación del fertilizante
fosfórico, así como de los fondos fijos de N y K2O, se tuvieron en cuenta los
requerimientos del cultivo y los tenores de estos nutrientes en el suelo, el
número de cortes por año y el rendimiento esperado. Se utilizó la cepa de HMA
INCAM-4 de la especie Glomus cubense (12), nombrada antes de su clasificación
como Glomus hoi-like, debido al alto índice de eficiencia que mostró en ensayos
realizados en condiciones similares a las que se condujo este experimento (13).
Para su aplicación se empleó un inoculante sólido certificado que contenía 25
esporas g-1 de sustrato, producido por el Departamento de Biofertilizantes
y Nutrición de las Plantas del INCA. El pasto se inoculó al momento de la siembra
por el método del recubrimiento de la semilla (14).
El primer corte se realizó a los 120 días después de la siembra y posteriormente
a intervalos de 45 y 90 días durante los períodos lluviosos y poco lluviosos,
respectivamente, para un total de seis cortes por año. Estos se realizaron a
una altura de 10 cm de la superficie del suelo y en cada uno se pesó la masa
verde (MV) de la parte aérea de las plantas ubicadas dentro del área de cálculo
de las parcelas, y se tomaron muestras de 200 g para determinar el porcentaje
de masa seca (MS) y las concentraciones de P de la biomasa aérea (11).
El rendimiento de MS y la extracción de P en la biomasa aérea se calcularon,
en el primer caso, a partir del rendimiento de MV y el por ciento de MS, y en
el segundo, a partir del rendimiento de MS y las concentraciones de P en la
biomasa aérea, respectivamente.
En cortes alternos, de cada parcela se tomaron 10 submuestras de raíces y de
suelo de la rizosfera a una profundidad de 0-20 cm, mediante el empleo de un
cilindro metálico de 5 cm de diámetro y 20 cm de altura. Los puntos de muestreo
se distribuyeron equidistantes y separados a 10 cm de los surcos (4).
Estas se homogenizaron para formar una muestra compuesta por parcela, de las
cuales se extrajo 1 g de raicillas para su tinción y clarificación (15). Se
evaluaron la colonización micorrízica (16), la densidad visual o intensidad
de la colonización (17) y el número de esporas en la rizosfera (18).
Los datos se procesaron mediante el análisis de varianza bifactorial, según
el diseño experimental utilizado, y la dócima de Duncan a p<0,05 (19) o el
intervalo de confianza a a = 0,05 (20), para establecer las diferencias significativas
entre las medias de los tratamientos. Se utilizó el programa estadístico SPSS
11.5 para Windows.
RESULTADOS
Y DISCUSIÓN
La figura
muestra el efecto de la inoculación de G. cubense, sola o combinada con las
aplicaciones de dosis crecientes de fertilizante fosfórico, en las variables
micorrízicas del pasto.
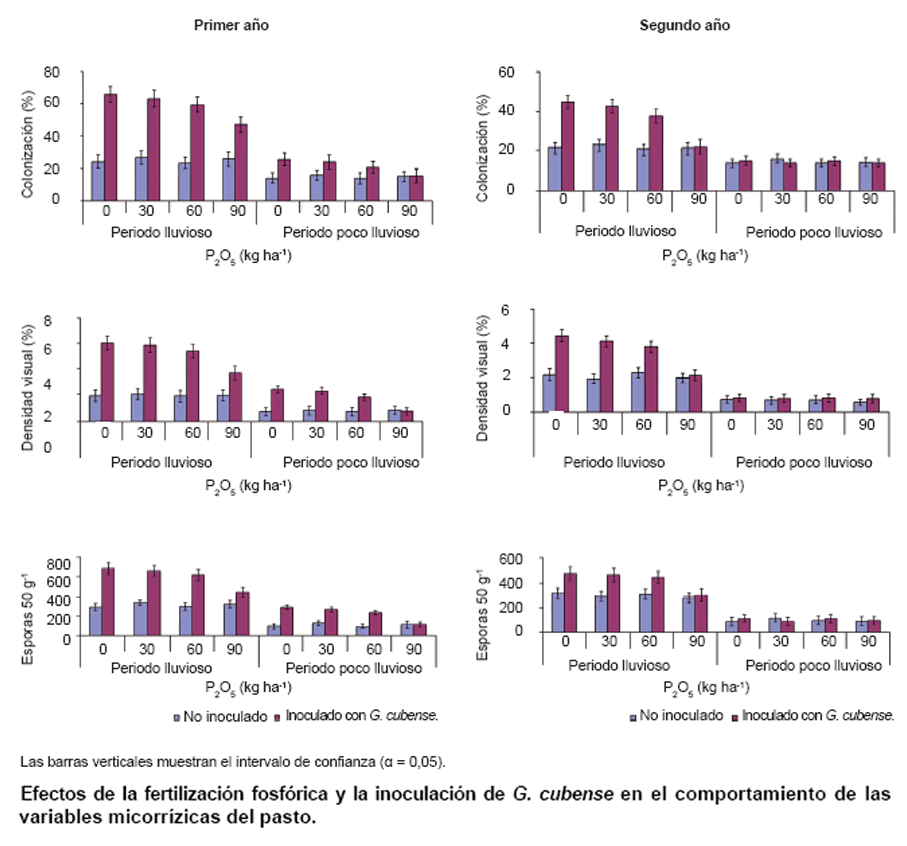
Se presentan los valores medios de cada variable durante los períodos lluvioso
y poco lluvioso de cada año. G. cubense incrementó el porcentaje de colonización,
la densidad visual y la densidad de esporas en la rizosfera, en relación con
los tratamientos no inoculados, los cuales exhibieron el nivel de ocupación
radical de los HMA residentes o nativos. La fertilización fosfórica no influyó
en estas variables; excepto en el tratamiento donde G. cubense se combinó con
la dosis más alta de P2O5, el cual presentó niveles de
colonización y densidad visual, así como cantidades de esporas significativamente
más bajos que el resto de los tratamientos inoculados.
El efecto de la inoculación de G. cubense permaneció hasta el período lluvioso
del segundo año, salvo en el tratamiento donde se aplicó la dosis más alta de
P2O5; no obstante, los valores absolutos de las variables
fúngicas fueron más bajos que durante el mismo período del año anterior. Los
porcentajes de colonización, densidad visual y la densidad de esporas en la
rizosfera fueron significativamente más altos en la época de lluvias que en
la época de seca de cada año.
Se encontró interacción significativa entre la fertilización fosfórica y la
inoculación o no de G. cubense para el rendimiento de MS del pasto (Tabla
II). En los tratamientos no inoculados, los mayores rendimientos se obtuvieron
con las dosis de 30 a 90 kg de P2O5 ha-1 año-1,
sin diferencias significativas entre ellas; sin embargo, sólo con la inoculación
de G. cubense, durante el primer año se obtuvieron valores similares a los alcanzados
con el fertilizante fosfórico sin inocular, de modo que las aplicaciones de
este nutriente resultaron innecesarias.
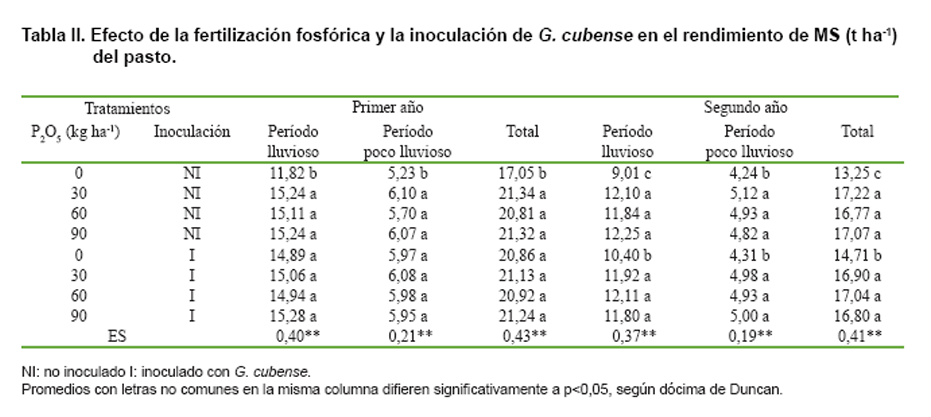
En el segundo año, la influencia de G. cubense en el rendimiento del pasto fue
menor que en el primero, pues solo se extendió hasta la época de lluvias y si
bien durante este período produjo valores significativamente mayores que los
alcanzados en el testigo sin P2O5 ni inoculación, su efecto
en la reducción de la respuesta del pasto al fertilizante fosfórico no se mantuvo,
de modo que fue necesario aplicar este nutriente para alcanzar los mayores rendimientos.
Ni las aplicaciones de P2O5 ni la inoculación de G. cubense
influyeron en las concentraciones de P en la biomasa de la parte aérea (Tabla
III), pero entre ambos factores se observó interacción significativa para
la extracción que de este elemento realizó la propia biomasa aérea del pasto
(Tabla IV).
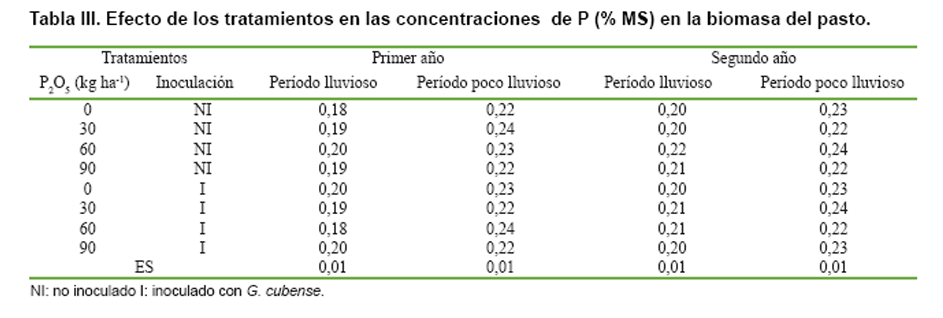
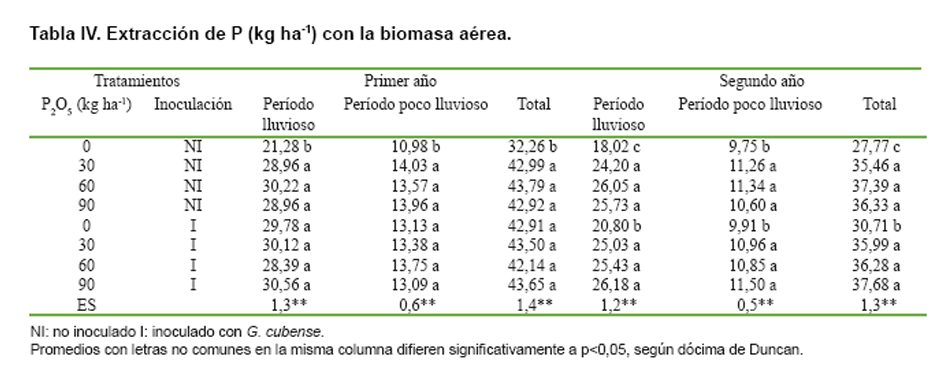
Como la extracción de P fue una función del rendimiento de MS del pasto, ambas
variables tuvieron un comportamiento muy similar; es decir, las mayores extracciones
de P se alcanzaron con las aplicaciones de de 30 a 90 kg de P2O5
ha-1 año-1 en los tratamientos no inoculados, sin diferencias
significativas con la producida durante el primer año sólo con la inoculación
de G. cubense. Durante el segundo año, el efecto de la cepa se mantuvo únicamente
hasta el período lluvioso, pero no alcanzó los niveles de extracción de P que
se obtuvieron con las aplicaciones del fertilizante fosfórico.
Varios aspectos deben resaltarse, a partir del análisis integral de los resultados
de este experimento. En primer lugar se demostró, en las condiciones en que
se realizó el presente estudio, la posibilidad de mejorar la nutrición fosfórica
y la productividad del pasto mediante la introducción de una cepa de HMA eficiente,
coincidiendo con los resultados de otros autores (21, 22), acerca de la contribución
de la inoculación de cepas de HMA eficientes al aumento de los rendimientos
y a la reducción los requerimientos de fertilizante fosfórico en especies forrajeras.
También se corroboró la efectividad de G. cubense para producir un funcionamiento
micorrízico adecuado y en consecuencia, una respuesta agronómica efectiva en
el pasto, aspectos comprobados anteriormente (12) en gramíneas forrajeras del
género Brachiaria y en otras plantas cultivadas en agroecosistemas con un tipo
de suelo similar al que se utilizó para experimento (23, 24).
En relación con el efecto de la inoculación de los HMA, aunque se reconoce que
la respuesta de los pastos suele variar en función de muchos factores debido
a que la ecofisiología de estos microorganismos en los agroecosistemas de pastizales
es bastante compleja (25), varios autores coinciden al plantear que la introducción
de cepas seleccionadas, adaptadas a las condiciones ambientales y con un alto
nivel de compatibilidad funcional y ecológica para el sistema suelo-planta,
puede ser una opción de manejo deseable e incluso necesaria, en los casos en
que las cepas residentes no sean lo suficientemente efectivas para producir
una respuesta agronómica deseada en estos cultivos (26).
La contribución de G. cubense a la nutrición fosfórica del pasto puede atribuirse
al aumento del coeficiente de aprovechamiento de este nutriente en virtud del
mayor volumen de suelo que pudieron explorar las raíces (27); ello se infiere
a partir de su efecto en las variables micorrízicas, cuyos incrementos, tanto
para la época lluviosa como para la poco lluviosa del primer año, parecen haber
contribuido decisivamente a la reducción de la necesidad de fertilizante fosfórico
durante ese período. De igual modo, la disminución que se observó en dichas
variables durante el segundo año debido a la reducción de la efectividad de
la cepa introducida, así como la necesidad de aplicar fertilizante fosfórico
para alcanzar los mayores rendimientos durante esa época, refuerzan esta hipótesis.
Además del efecto físico de G. cubense en la extensión del sistema radical,
otros mecanismos relacionados con la capacidad de los HMA para acceder incluso
a fuentes de P menos disponibles y facilitar su absorción por las plantas, también
pudieran explicar su influencia en la nutrición fosfórica del pasto (28, 29,
30).
Por otra parte, cuando la inoculación de G. cubense se combinó con la dosis
más alta de P2O5 se observó una reducción espacial y temporal
de las variables fúngicas, lo que parece ser una consecuencia fisiológica de
la disminución del papel de las micorrizas en la absorción de este elemento,
debido a la aplicación de una cantidad de fertilizante fosfórico que evidentemente
resultó mucho mayor que la requerida por el pasto.
En tal sentido, algunos autores plantean que la disponibilidad de nutrientes
controla el crecimiento de las estructuras micorrízicas y que la distribución
de arbúsculos e hifas extrarradicales se reduce cuando los pastos han sido suficientemente
fertilizados, ya que la entrega de los recursos del suelo a la planta hospedera
a través de los HMA pierde importancia (31, 32).
Llama la atención el hecho de que la aplicación de una dosis baja de fósforo
(30 ka de P2O5 ha-1 año-1) haya
sido suficiente para obtener el máximo rendimiento del pasto no inoculado, a
pesar del bajo contenido inicial de P2O5 asimilable del
suelo (9). Asimismo, el hecho de que ni la fertilización fosfórica ni la inoculación
de G. cubense hayan incrementado los tenores de P en la biomasa, también parece
corroborar el bajo requerimiento externo de P del pasto. En este sentido, se
ha podido comprobar que las raíces de B. decumbens pueden acceder a formas menos
disponibles de P en el suelo (33, 34).
El hecho de que con la inoculación de una cepa de HMA eficiente se haya podido
prescindir de la fertilización fosfórica para mejorar el rendimiento del pasto,
al menos durante el primer año de su establecimiento, indica que además de las
características morfológicas de la raíz y los mecanismos bioquímicos que pueden
estar involucrados en la baja respuesta de esta especie a las aplicaciones de
P, G. cubense jugó un papel decisivo en el mejoramiento de la nutrición fosfórica
de este cultivo y de hecho en la reducción de los requerimientos externos de
este elemento.
No menos interesantes resultaron las variaciones estacionales que presentaron
las variables micorrízicas, pues durante los dos años en todos los tratamientos
se observaron niveles de colonización, densidad visual y densidad de esporas
significativamente mayores en el período lluvioso que en el poco lluvioso y
ello coincidió con la distribución de la producción de biomasa del pasto en
una y otra épocas.
En este sentido, algunos autores plantean que el crecimiento acelerado de los
pastos durante el período en que concurren los mayores niveles de precipitaciones,
temperatura y luminosidad, demanda una mayor cantidad de nutrientes y agua y
por lo tanto, mayores cantidades de estructuras micorrízicas para acceder a
tales recursos (35, 36). Además, como el costo de la colonización micorrízica
suele ser alto (se plantea que hasta un 20 % del C fijado puede ser entregado
al simbionte microbiano), durante la época de mayor crecimiento las plantas
estarían en mejores condiciones de destinar recursos para la formación y el
mantenimiento de dichas estructuras (37).
CONCLUSIONES
A partir de estos
resultados se puede concluir que la inoculación de una cepa de HMA eficiente
incrementa las estructuras micorrízicas y reduce la necesidad de aplicar fertilizante
fosfórico al pasto B. decumbens, cv. Basilisk, al menos durante el primer año
de su establecimiento. De este modo, el manejo de los HMA se vislumbra como
una alternativa económica y ecológicamente viable para disminuir los costos
de la fertilización de los pastos y los riesgos de contaminación ambiental que
conlleva su aplicación extensiva y sistemática a estos cultivos.
BIBLIOGRAFÍA
1. Santos, M.
E. R.; da Fonseca, D. M.; Balbino, E. M.; Monnerat, J. P. I. S. y da Silva,
S. P. Capim-braquiária diferido e adubado com nitrogênio: produção e características
da forragem. R. Bras. Zootec., 2009, vol. 38, no. 4, pp. 650-656. ISSN 1806-9290.
2. Martha Jr., G. B.; Corsi, M.; Trivelin, P. C. O. y Vilela, L. R. Recuperação
de 15N-ureia no sistema solo-planta de pastagem de capim-tanzânia. R. Bras.
Ci. Solo., 2009, vol. 33, pp. 95-101. ISSN 0100-0683.
3. Richardson, A. E.; Linch, J. P.; Ryan, P. R.; Delhaize, E.; Smith, F. A.;
Smith S. E.; Harvey, P. R.; Ryan, M. H.; Veneklaas, E. J.; Lambers, H.; Oberson,
A.; Culvenor, R. A. y Simpson, R. J. Plant and microbial strategies to improve
the phosphorus efficiency of agriculture. Plant and Soil., 2009, vol. 349, pp.
121-156. ISSN 0032-079X.
4. Johnson, N. C.; Rowland, D. L.; Corkidi, L.; Egerton, L. M. y Allen, E. B.
Nitrogen enrichment alters mycorrhizal allocation at five mesic to semi-arid
grasslands. Ecology., 2003, vol. 84, no. 7, pp. 1895-1908. ISSN 0012-9658.
5. Leigh, J.; Hodge, A. y Fitter, A. H. Arbuscular mycorrhizal fungi can transfer
substantial amounts of nitrogen to their host plant from organic material. New
Phytologist., 2009 no. 181, pp. 199-207. ISSN 1469-8137.
6. Hu, J.; Lin, X.; Wang, J.; Dai, J.; Cui, X.; Chen, R. y Zhang, J. Arbuscular
mycorrhizal fungus enhances crop yield and P-uptake of maize (Zea mays L.):
A field case study on a sandy loam soil as affected by long-term P-deficiency
fertilization. Soil Biology & Biochem., 2009, vol. 41, pp. 2460-2465. ISSN
0038-0717.
7. Costa, K. A.; Severiano, E. C.; Simon, G. A. y Carrijo, M. S. Extração de
nutrientes do capim-marandu sob doses e fontes de nitrogênio. Rev. Bras. Saúde
Prod. Anim., 2009, vol. 10, no. 4, pp. 801-812. ISSN 1519-9940.
8. Ramos, S. J.; Faquin, V.; Rodrigues, C. R. Silva, C, A. y Boldrin, P. F.
Biomass production and phosphorus use of forage grasses fertilized with two
phosphorus sources. R. Bras. Ci. Solo., 2009, vol.33, pp. 335-343. ISSN 0100-0683.
9. Hernández, A.; Pérez, J. M.; Bosch, D. y Rivero, L. Nueva Versión de clasificación
genética de los suelos de Cuba. La Habana: AGRINFOR. 1999. 64 pp. ISBN 959-246-022.
10. Paneque, V. M. y Calaña, J. M. La fertilización de los cultivos. Aspectos
teórico prácticos para su recomendación. Departamento de Biofertilizantes y
Nutrición de las Plantas. La Habana: INCA. 2001. 29 pp.
11. Paneque, V. M.; Calaña, J. M.; Calderón, M.; Borges, Y.; Hernández, T. y
Caruncho. M. Manual de técnicas analíticas para análisis de suelo, foliar, abonos
orgánicos y fertilizantes químicos. Ediciones INCA, San José de las Lajas, La
Habana, Cuba, 2010, 160p. [Consultado: 21 may 2011]. Disponible en: <http://ediciones.inca.edu.cu/files/folletos/folleto_suelos.pdf>.
ISBN 978-959-7023-51-7.
12. Rodríguez, Y.; Dalpé, Y.; Séguin, S; Fernández, K.; Fernández, F. y Rivera,
R. A. Glomus cubense sp. nov., an arbuscular mycorrhizal fungus from Cuba. Micotaxon,
2011, vol. 118, pp. 337-347. ISSN 0187-3180.
13. González, P. J.; Arzola, J.; Morgan, O.; Rivera, R.; Plana, R. y Fernández,
F. Manejo de las asociaciones micorrízicas en pastos del género Brachiaria cultivados
en suelos Ferralítico Rojo y Pardo Mullido. En Congreso Científico del INCA
(16:2008, nov 24 - 28, La Habana). Memorias. CD-ROM. Instituto Nacional de Ciencias
Agrícolas. ISBN 978-959-16-0953-3.
14. Fernández, F.; Gómez, R.; Vanegas, L. F.; Martínez, M. A.; de la Noval,
B. M. y Rivera, R. Producto inoculante micorrizógeno. Oficina Nacional de Propiedad
Industrial. Cuba, Patente No. 22641, 2000.
15. Phillips, D. M. y Hayman, D. S. Improved procedures for clearing roots and
staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment
of infection. Trans. Br. Mycol. Soc., 1970, vol. 55, pp. 158-161. ISSN 0007-1536.
16. Giovanetti, M. y Mosse, B. An evaluation of techniques for measuring vesicular-
arbuscular mycorrhizal infection in roots. New Phytol., 1980, vol. 84, pp. 489-500.
ISSN 1469-8137.
17. Trouvelot, A.; Kough, J. y Gianinazzi-Pearson, V. Mesure du taux de mycorhization
VA d'un systeme radiculaire. Recherche de methodes d'estimation ayantune signification
fonctionnelle. En: Proc. 1st European. Symposium on Mycorrhizae: Physiological
and genetical aspects of mycorrhizae, (1986, jul. 1-5: Dijón), 1986. pp. 217-222.
18. Gerdemann J. W. y Nicolson, T. H. Spores of mycorrhizal Endogone species
extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc., 1963,
vol. 46, pp. 235-244. ISSN 0007-1536.
19. Duncan, D. B. Multiple range and multiple F test. Biometrics, 1955, vol.
11, pp. 1-42. ISSN 1541-0420.
20. Payton, M.
E.; Miller, A. E. y Raun, W. R. Testing statistical hypotheses using standard
error bars and confidence intervals. Comm. Soil Sci. Plant Anal., 2000, vol.
31, pp. 547-551. ISSN 1532-2416.
21. Carneiro, R. F. V.; Martins, M. A.; Vásquez, H. M. y Detmann, E. Doses de
fósforo e inoculação micorrízica no cultivo de estilosantes em solo sob condições
naturais. Arch. Zootec., 2010, vol. 59, no. 227, pp. 415-426. ISSN 0004-0592.
22. Carneiro,
R. F. V.; Martins, M. A.; Freitas, Marta, S. M.; Detmann E. y Vasquez, H. M.
Inoculação micorrízica arbuscular e doses de fósforo na produção do capim-andropogon,
em substrato não estéril. Rev. Bras. Cienc. Agr., 2007, vol. 2, no. 3, pp. 212-218.
ISSN 1981-0997.
23. Plana, R.; González, P. J.; Dell'Amico, J. M.; Fernández, F.; Calderón,
A. y Marrero, Y. Efecto de dos inoculantes micorrizicos arbusculares (base líquida
y sólida) en el cultivo del trigo duro (Triticum durum). Cultivos Tropicales,
2008, vol. 29, no. 4, pp.35-40. ISSN 1819-4087.
24. Martín, G.; Arias, L. y Rivera, R. Selección de las cepas de hongos micorrízicos
arbusculares (HMA) más efectivas para la Canavalia ensiformis cultivada en suelo
Ferralítico Rojo. Cultivos Tropicales, 2010, vol. 31, no. 1, pp. 27-31. ISSN
1819-4087.
25. Bingham, M. A. y Biondini, M. Mycorrhizal hyphal length as a function of
plant community richness and composition in restored northern tallgrass prairies
(USA). Rangeland Ecol. Manag., 2009, vol. 62, pp. 60-67. ISSN 1551-5028.
26. Miranda de, E. M.; Júnior. Seleção de fungos micorrízicos arbusculares para
o amendoim forrageiro consorciado com braquiária. Pesq. Agropec. Bras. 2008,
vol. 43, no. 9, pp. 1185-1191. ISSN 1678-3921.
27. Richardson, A. E.; Hocking, P. J.; Simpson, R. J. y George, T. S. Plant
mechanisms to optimise access to soil phosphorus. Crop & Pasture Science.,
2009, vol. 60, pp. 124-143. ISSN 1836-0947.
28. Yao, Q.; Zhu, H. H.; Hu, Y. L. y Li, L. Q. Differential influence of native
and introduced arbuscular mycorrhizal fungi on growth of dominant and subordinate
plants. Plant Ecol., 2008, vol.196, pp. 261-268. ISSN 1573-5052.
29. Balota, E. L.; Machineski, O. y Stenzel, N. M. C. Resposta da acerola à
inoculação de fungos micorrízicos arbusculares em solo com diferentes níveis
de fósforo. Bragantia, 2011, vol. 70, no. 1, pp. 166-175. ISSN 1678-4499.
30. Almagrabi, O. A. y Abdelmoneim T. S. Using of arbuscular mycorrhizal fungi
to reduce the deficiency effect of phosphorous fertilization on maize plants
(Zea mays L.). Life Science Journal, 2012, vol. 9, pp. 1649-654. ISSN 1097-8135.
31. Rai, A.; Rai, S. y Rakshit, A. Mycorrhiza-mediated phosphorus use efficiency
in plants. Environm. Exp. Biol., 2013, vol. 11, pp. 107–117. ISSN 2255-9582.
32. Netu, N.; Aggarwal, A.; Tanwar, A. y Alpa, A. Influence of arbuscular mycorrhizal
fungi and Pseudomonas fluorescens at different superphosphate levels on linseed
(Linum usitatissimum L.) Growth response. Chilean J. of Agric. Res., 2012, vol.
72, pp. 237-243. ISSN 0718-5839.
33. Magalhaes, A. F.; Pires, A. J. V.; Carvalho, G. G. P.; Silva, F. F.; Sousa,
R. S. y Veloso, C. M. Influência do nitrogênio e do fósforo na produção do capim-braquiária.
Revista Brasileira de Zootecnia, 2007, vol. 36, no. 5, pp. 1240-1246. ISSN 1806-9290.
34. Low-Gaume, A. E. Morphological, physiological and biochemical adaptation
of Brachiaria grasses to low phosphorus supply. Disertación para obtener el
grado de Dr. C. Federal Institute of Technology (ETH). Swiss Centre for International
Agriculture, Disert. No. 18131. ETH, Zürich, 2009.
35. García, I. V. y Mendoza, R. E. Relationships among soil properties, plant
nutrition and arbuscular mycorrhizal fungi-plant symbioses in a temperate grassland
along hydrologic, saline and sodic gradients. FEMS Microbiology Ecology, 2008,
vol. 63, pp. 359. ISSN 1574-6941.
36. Mandyam, K. y Jumpponen, A. Seasonal and temporal dynamics of arbuscular
mycorrhizal and dark septate endophytic fungi in a tallgrass prairie ecosystem
are minimally affected by nitrogen enrichment. Mycorrhiza, 2008, vol. 18, pp.
145-155. ISSN 1432-1890.
37. Siddiqui, Z. A.; Sayeed, M. y Futai, K. Mycorrhizae: sustainable agriculture
and forestry. Ed: Springer Science + Business Media B. V., 2008, 359 pp. ISBN
978-1-4020-8769-1.
Recibido: 4 de octubre de 2013
Aceptado: 11 de diciembre de 2014
Ms.C. Pedro
J. González Cañizares, Instituto Nacional de Ciencias Agrícolas (INCA),
gaveta postal 1, San José de las Lajas, Mayabeque. Cuba. CP 32 700. Email: pgonzalez@inca.edu.cu