Análisis de la interacción progenie-ambiente con minitubérculos a partir de semilla sexual de papa (Solanum tuberosum, L.) en Cuba
Analysis of progeny-environment interaction with seedling tubers of true potato seed (Solanum tuberosum L.) in Cuba
Ms.C. J. L. Salomón,I Dr.C. J. G. Castillo,I Dr.C. J. A. Arzuaga,I Dr.C. W. Torres,I Dr.C. A. Caballero,I Dr.C. M. Varela,I Victor Manuel Hernández BetancourtII
IInstituto
Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José
de las Lajas, Mayabeque, Cuba, CP 32700.
IIFacultad
Agropecuaria de Montaña del Escambray (FAME) Tope de Collantes, Trinidad,
Universidad de Sancti Spiritus, UNISS.
RESUMEN
Durante las campañas agrícolas 2007-2008 y 2008-2009 en tres localidades (Instituto Nacional de Ciencias Agrícolas-Mayabeque, Empresa Agropecuaria “V. Lenin”-Matanzas y Empresa Agropecuaria “La Cuba”-Ciego de Ávila), se evaluaron 13 progenies de semilla sexual de papa y tres cultivares testigos (Romano, Spunta y Cal White). Se registraron número de tubérculos por planta, rendimiento por hectárea (t ha-1) e incidencia de manchas en el follaje del hongo Alternaria solani Soraeur. Los datos fueron procesados mediante un análisis de varianza de clasificación doble, y se ajustó el modelo de efectos principales aditivos e interacciones multiplicativas (AMMI) a los caracteres donde resultó significativa la interacción genotipo-ambiente para el ajuste. La progenie Lajera x 6-3-98 fue estable para el carácter número de tubérculo y rendimiento, en los seis ambientes estudiados. Las progenies Lajera x 6-3-98, Lajera x 9-80-98, fueron estables para el rendimiento y para la masa promedio del tubérculo en los seis ambientes. Las progenies Samila x 2-130-98, Yara x 1-10-96, Yara x 9-80-98, Atlantic x Aninca y Samila x 6-3-98, fueron las más inestables para las características estudiadas pero con adaptación específica en cada localidad. Estos contrastes en el comportamiento agronómico de las progenies de semilla sexual sugiere la necesidad de evaluar los materiales en cada zona de producción..
Palabras clave: papa, progenie sexual, cultivar, ambiente, estabilidad, rendimiento.
ABSTRACT
During crop years 2007-2008 and 2008-2009 at three locations ( National Institute of Agricultural Sciences-Mayabeque , Agricultural Enterprise “V. Lenin”-Matanzas and Agricultural Enterprise “La Cuba”-Ciego de Ávila), 13 progenies from seedling tubers were evaluated and three witnesses varieties (Romano, Spunta and Cal White). Number of tubers per plant, yield per hectare (t ha-1) and incidence of spots on the foliage by the fungus Alternaria solani Soraeur were recorded. The data were processed by analysis of variance, and adjusted with additive main effects and multiplicative interaction model (AMMI) to the characters which was significant genotype environment interaction was adjusted. The sexual progeny Lajera x 6-3-98 were stable for tuber number and yield in six environments studied. The Lajera x 6-3-98, Lajera x 9-80-98, progenies were stable for yield and average tuber mass in the six environments studied. The progeny Samila x 2-130-98, Yara x 1-10-96, Yara x 9-80-98, Atlantic x Aninca y Samila x 6-3-98, were the most unstable for the three studied traits but specific adaptation in each locality. These contrasts in the agronomic performance of the progeny of sexual seed suggest the need to evaluate the materials in each production area.
Key words: potato, sexuwal progeny, cultivar, environment, stability, yield.
INTRODUCCIÓN
La papa (Solanum
tuberosum) es una planta perteneciente al género Solanum, originaria
de América del Sur y cultivada en todo el mundo por sus tubérculos
comestibles. Es uno de los cultivos alimenticios básicos que más
expande su superficie de plantación en los países en desarrollo,
esto se debe a su amplia adaptación, a su alto rendimiento por unidad
de superficie y de tiempo, a su gran valor nutritivo y a las variadas formas
de uso y de productos procesados que de ella se obtienen (1).
Este cultivo se reproduce comercialmente mediante tubérculos-semilla;
sin embargo, también puede hacerlo por vía sexual con semilla
verdadera o botánica (SB), lo que es aprovechado, principalmente, para
el mejoramiento genético de la especie, pero también, como material
de siembra para producir papa semilla y papa consumo (2).
Los tubérculos-semilla por lo general son el insumo más costoso
en la producción de papa, y en Cuba representan entre el 40 y el 50 %
(1200-1500 USD ha-1) de los mismos, teniendo en cuenta el precio de la SB en
el mercado internacional, se puede reducir por concepto de semilla entre el
80 y el 90 % por hectárea.
El cultivo está influenciado por elementos meteorológicos que
influyen sobre el crecimiento, desarrollo, producción y calidad de la
papa. Estos son principalmente la temperatura del suelo y el aire, radiación
solar, fotoperíodo, humedad del suelo y evapotranspiración (3).
La evaluación de los genotipos en diferentes localidades y a través
del tiempo es importante para estimar las respuestas fenotípicas y genotípicas
diferenciales bajo diversas condiciones ambientales; medir su interacción
genotipo-ambiente da una idea de la estabilidad fenotípica de las variedades
ante las fluctuaciones ambientales (4), este análisis ha sido utilizado
con éxito para seleccionar genotipos de papa deseables (5).
La papa, mediante la reproducción sexual produce una descendencia altamente
heterogénea, ya que los clones son heterocigotas, como resultado, cada
semilla será genéticamente diferente y por consiguiente, la producción
de tubérculos también poseerá alta variabilidad. Esto sería
un obstáculo para el empleo de la tecnología de SB, pero otorgaría
una mayor estabilidad y adaptación de estas a diferentes ambientes.
Teniendo en cuenta los argumentos antes señalados el objetivo de este
trabajo fue evaluar el comportamiento agronómico, la estabilidad y adaptabilidad
de progenies de SB de papa en las tres localidades más importantes para
el cultivo, durante dos años y seleccionar materiales en relación
a la interacción genotipo-ambiente.
MATERIALES Y MÉTODOS
Durante las campañas
agrícolas 2008-2009 y 2009-2010 se evaluaron en tres localidades (Instituto
Nacional de Ciencias Agrícolas “INCA”-Mayabeque, Empresa Agropecuaria
“Lenin”- Matanzas y Empresa Agropecuaria “La Cuba”- Ciego de Ávila) 13
progenies de SB (9-80-98-PL; Atlantic x 6-3-98; Atlantic x Aninca; Gorbea x
1-10-98; Lajera x 6-3-98; Gorbea x 6-3-98; Lajera x 9-80-98; Samila x 2-130-98;
Samila x 6-3-98; Samila-PL; Yara x 1-10-98; Yara x 9-80-98; Yara-PL y tres cultivares
testigos (Cal White, Romano y Spunta).
Se utilizaron como material de plantación minitubérculos F1 obtenidos
en canteros de las progenies y tubérculos-semilla certificados calibre
28-35 mm para los cultivares testigo.
La fecha de plantación en cada localidad para la primera y segunda campaña
fueron 12, 15 y 17 de diciembre y 10, 12 y 15 de diciembre, respectivamente.
En todas las localidades los experimentos se llevaron a cabo en un diseño
de bloques completos al azar con cuatro réplicas. Cada parcela midió
2,25 x 1,8 m con dos surcos separados a 90 cm entre ellas y 25 cm entre plantas.
Se adoptaron las labores culturales recomendadas para el manejo del cultivo
(6).
En cada localidad fueron registrados los siguientes datos:
1. Número de tubérculos por planta: se cosecharon cinco plantas
seleccionadas al azar en cada parcela y se les contó el número
de tubérculos.
2. Rendimiento (t ha-1): rendimiento (kg) por planta por el total
de plantas en una hectárea (44444) y dividido por 1000.
3. Incidencia de manchas en el follaje de infección por el hongo Alternaria
solani (Ellis y Martín) Jones y Grout, se realizó a los 65
días de la plantación, para la cual se utilizó la escala
de 9 grados propuesta por Horsfall y Barrat citados por Estévez (7),
donde el grado 1- sin manchas en el follaje y grado 9- todas las hojas del follaje
muertas.
Análisis estadístico
Los datos fueron procesados a través de un análisis de varianza
de clasificación doble y se le aplicó la prueba de rango múltiple
de Duncan al 5 % mediante el paquete estadístico SPSS (16.0). Para los
caracteres donde se detectó una interacción genotipo-ambiente
significativa, se analizó la misma por medio del modelo AMMI, se ajustó
el modelo AMMI con efecto interacción multiplicativo y efectos principales
aditivos (8), para el cual se utilizó el programa SAS.
RESULTADO Y DISCUSIÓN
Al realizar el
análisis de varianza correspondiente, se obtuvo una interacción
entre progenies y entre progenie x ambiente significativas para el carácter
número de tubérculos por planta, rendimiento total (t ha-1)
y masa promedio del tubérculo, lo que justificó el ajuste del
modelo AMMI.
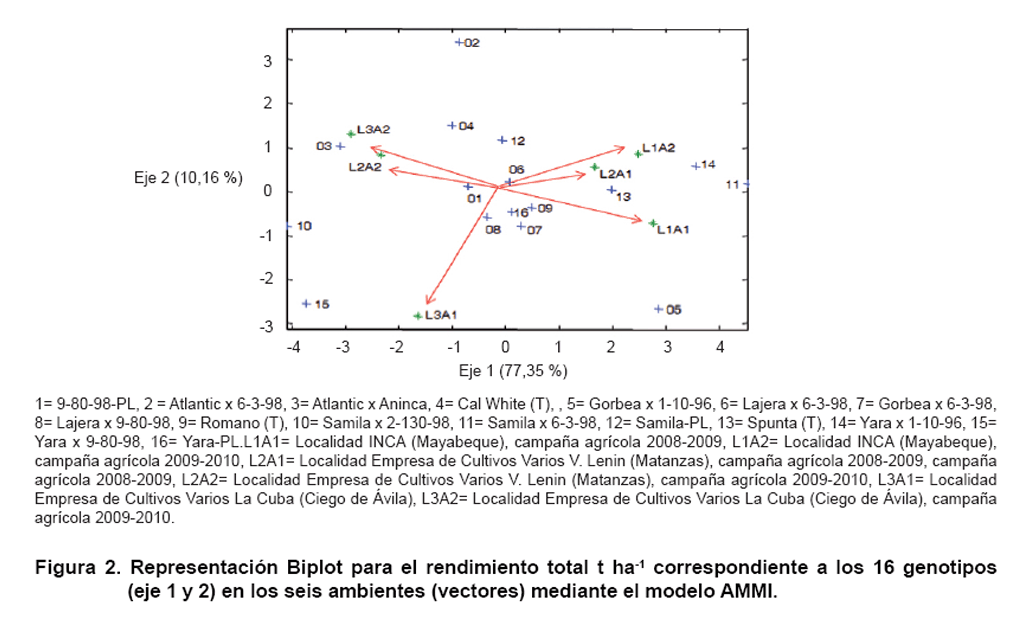
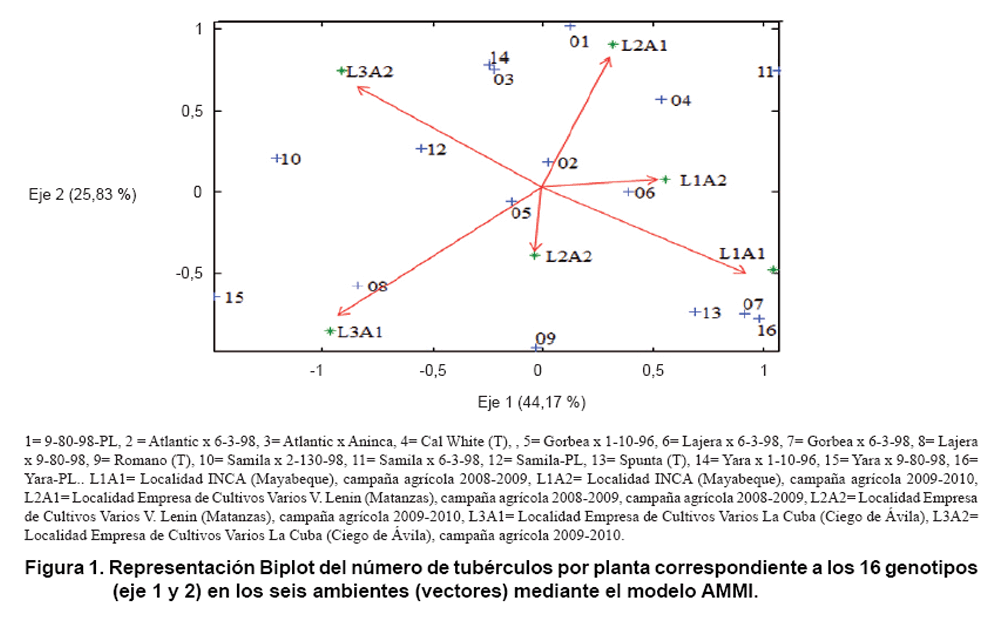
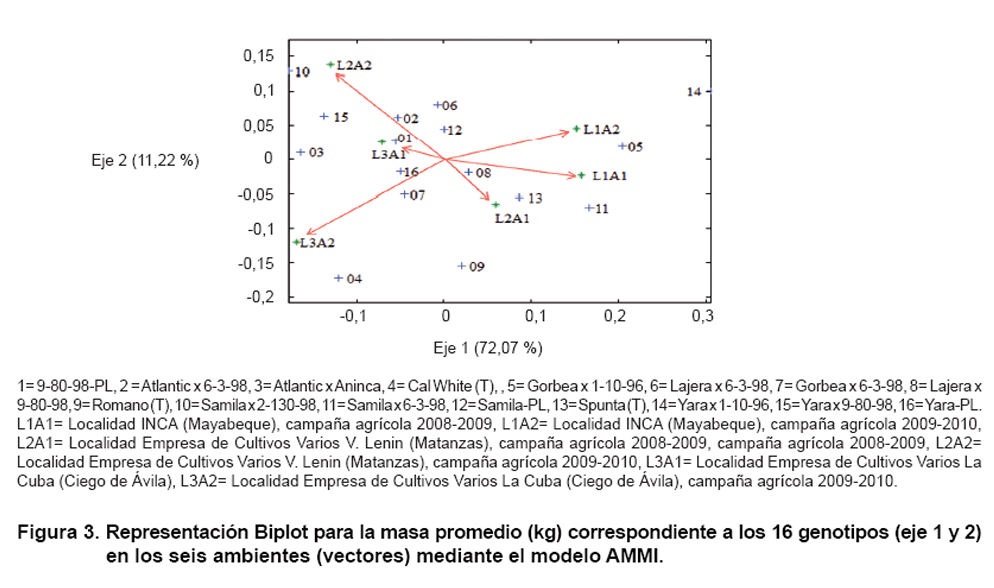
En la gráfica Biplot (Figura
1, 2 y 3)
los genotipos fueron representados por un punto y los ambientes, como vectores
con origen en el punto 0,0. Mientras más cerca se localizan los genotipos
al centro de la gráfica, menos responden a las variaciones ambientales,
por tanto, son los más estables. Entre tanto, los más alejados
del centro son considerados los que más aportan a la interacción,
o sea, que más responden a los estímulos ambientales.
En la Figura 1, ambos
ejes (1 y 2) explicaron el 70,0 % de la interacción genotipo-ambiente
para el número de tubérculos por planta, el eje 1 explica el 44,17
% de la interacción, mientras el eje 2, los restantes 25,83 %.
Las progenies 2 (Atlantic x 6-3-98), 5 (Gorbea x 6-3-98) y 6 (Lajera x 6-3-98)
fueron las más estables para este carácter en los seis ambientes
estudiados.
Se observó en esta misma figura, una respuesta positiva a los estímulos
ambientales en L3A1, a las progenies 8 (Lajera x 9-80-98), 15 (Yara x 9-80-98)
y el cultivar testigo 9 (Romano) y negativamente en L2A1. Por otro lado, las
progenies 7 (Gorbea x 6-3-98), 16 (Yara- PL) y el cultivar testigo 13 (Spunta)
interactuaron positivamente en el ambiente L1A1, pero de forma negativa en L2A1
y L3A2. También respondieron positivamente las progenies 1 (9-80-98-PL),
3 (Atlantic x Aninca), el cultivar testigo 4 (Cal White), 11 (Samila x 6-3-98)
y 14 (Yara x 1-10-98) pero en el ambiente L2A1 y de forma negativa en L3A1.
Finalmente la progenie 10 (Samila x 2-130-98) interactuó positivamente
en el ambiente L3A2 y negativamente en L1A1.
Los resultados sugieren la capacidad de un grupo de progenies con adaptación
a todos los ambientes estudiados y otro grupo de adaptarse a medios específicos.
Estos resultados indican que la prueba de progenie en múltiples localidades
puede identificar progenitores que producen progenies más ampliamente
adaptadas, y por otro lado, seleccionar progenies superiores con adaptación
general y específica.
Resultados similares indicó Sharma et al. (9), quienes establecieron
que las localidades, los genotipos y la interacción localidad x genotipo
mostraron variaciones significativas para el número de minitubérculos
por planta en dos épocas de cultivo (óptima y tardía) en
el Himalaya.
En la Figura 2, ambos
ejes (1 y 2) explicaron el 87,51 % de la interacción genotipo-ambiente
para el rendimiento (t ha-1), el eje 1 explica el 77,35 % de la interacción,
mientras el eje 2, los restantes 10,16 %.
Respondieron positivamente en los ambientes L2A2 y L3A2, las progenies 3 (Atlantic
x Aninca), la 10 (Samila x 2-130-98) y la 2 (Atlantic x 6-3-98), pero de forma
negativa en L1A1 y L1A2 a excepción de la progenie 2 que lo hizo en L3A1.
Entre tanto, la 15 (Yara x 9-80-98) interactuó positivamente en L3A1
y negativamente en L1A2. Las progenies 11 (Samila x 6-3-98) y 14 (Yara x 1-10-96)
tuvieron una respuesta positiva en L1A2 y L1A1 y respondieron negativamente
en L3A2 y L2A2. La progenie 5 (Gorbea x 1-10-96) interactuó positivamente
en L1A1 y negativamente en L3A2 y L2A2.
Por otro lado, las progenies 1 (9-80-98-PL), 6 (Lajera x 6-3-98), 7 (Gorbea
x 6-3-98), 8 (Lajera x 9-80-98), 16 (Yara-PL) y el cultivar testigo 9 (Romano)
fueron las más estables en el rendimiento, lo cual podría estar
indicando que estos genotipos portan genes estabilidad en estos medios, los
cuales pueden estar presentes, independientemente del origen del genotipo (PL,
híbridos o cultivares). Este resultado podría indicar que el mismo
efecto ambiental no actúa en igual medida sobre el comportamiento de
las progenies, las mismas mostraron comportamiento diferenciado, estudio comparable
con lo informado por Navarro (10), quién indicó que los genes
no pueden hacer que se exprese un carácter si no tiene el adecuado y,
viceversa, ninguna manipulación ambiental hará que se desarrolle
una cierta característica si no están presentes los genes necesarios.
Estos resultados podrían estar expresando un mejor comportamiento de
las progenies híbridas y de PL que los genotipos testigos, los cuales
son cultivares comerciales con estabilidad y adaptación a ambientes contrastantes.
Los resultados en este trabajo sugieren la gran influencia que ejerce el medio
y que conlleva el comportamiento diferencial de los genotipos.
El rendimiento es un atributo altamente variable que está influenciado
por el año, la localidad, el tipo de suelo, la fertilidad del suelo,
la tecnología y el genotipo, por tanto, la estabilidad de un genotipo
en un amplio rango de ambientes tiene una importancia considerable en la selección
de progenitores y cultivares comerciales.
Los resultados alcanzados son similares a los obtenidos en otros estudios (7),
donde se informó que las variedades Romano y Desirée han sido
las más estables y de las de mejor comportamiento en la producción
durante varios años en Cuba. Los mismos coinciden con los encontrados
por otros autores (11), quienes determinaron los grandes contrastes de rendimiento
de tubérculos a partir de semilla botánica entre diferentes localidades,
lo que se atribuye a la interacción genotipo-ambiente, y sugiere la necesidad
de evaluar los genotipos en cada zona de producción (12).
Se ha reportado la utilización del modelo de efectos principales aditivos
e interacción multiplicativa (AMMI) para clasificar los ambientes con
los mayores rendimientos de tubérculo, y seleccionar los genotipos más
sobresalientes y con correlación positiva con dos localidades y dos años.
En este mismo sentido, algunos autores establecieron, mediante un modelo AMMI,
las variedades de papa más estables para caracteres del rendimiento y
sus componentes en nueve localidades diferentes (8).
Aunque el rendimiento es el resultado de los efectos del genotipo (G), del ambiente
(E) y de la interacción genotipo-ambiente (GE), solamente el efecto principal
de G y la interacción GE son de relevancia en la evaluación de
los genotipos (13).
La diferenciación de medios permitió identificar comportamientos
diferenciados entre los genotipos, los cuales podrían ser utilizados
en un programa de mejoramiento genético o seleccionarlos para ser empleados
comercialmente, coincidiendo con lo recomendado cuando se analizó la
interacción genotipo x ambiente en papa utilizando la metodología
AMMI para la identificación de genotipos estables y adaptados a localidades
específicas (3).
En la Figura 3 ambos
ejes (1 y 2) están explicando el 83,29 % de la interacción genotipo-ambiente
para la masa promedio del tubérculo, el eje 1 explica el 72,07 % de la
interacción, mientras el eje 2 los restantes 11,22 %.
Las progenies 1 (9-80-98-PL), 7 (Gorbea x 6-3-98), 8 (Lajera x 9-80-98), 16
(Yara-PL) y 12 (Samila-PL) fueron las más estables para la masa promedio
del tubérculo.
En la Figura 3 se observa
que las progenies 10 (Samila x 2-130-98) y 15 (Yara x 9-80-98) interactuaron
de forma positiva en L2A2 y negativamente en L1A2 y L1A1. El cultivar testigo
4 (Cal White) interactuó positivamente en L3A2 y negativamente en L1A2
y L2A2. Mientras que en L1A2 y L1A1, respondieron positivamente las progenies
5 (Gorbea x 1-10-96) y 14 (Yara x 1-10-96), pero tuvieron una respuesta negativa
en L3A2.
Así mismo, recientemente se ha establecido un método efectivo
para estudiar la estabilidad de las variedades de papa en varias localidades
y años, utilizando un modelo AMMI generalizado y que pueden ser representados
mediante un Biplot interactivo (8).
En la Figura 4 se observa
de forma general, que las progenies híbridas y las variedades testigos
presentaron los menores grados de afectación de Alternaria solani,
agente causal del tizón temprano en el follaje, enfermedad importante
del cultivo de papa en Cuba. Estos resultados pudieran indicar que uno de los
criterios de selección empleado en el programa de mejoramiento genético
para la obtención de variedades, fue la tolerancia a esta enfermedad
en condiciones de campo. La misma se presenta todos los años y con variada
agresividad, debido probablemente a las condiciones climáticas y manejo
del cultivo, por lo cual se ha convertido en una patología clave del
cultivo que, potencialmente, puede provocar importantes pérdidas, resultado
que corrobora lo determinado por algunos autores (14), según lo cual
esta enfermedad se presenta prácticamente todos los años y con
variada intensidad, debido probablemente, a la gran amplitud de los momentos
de plantación.
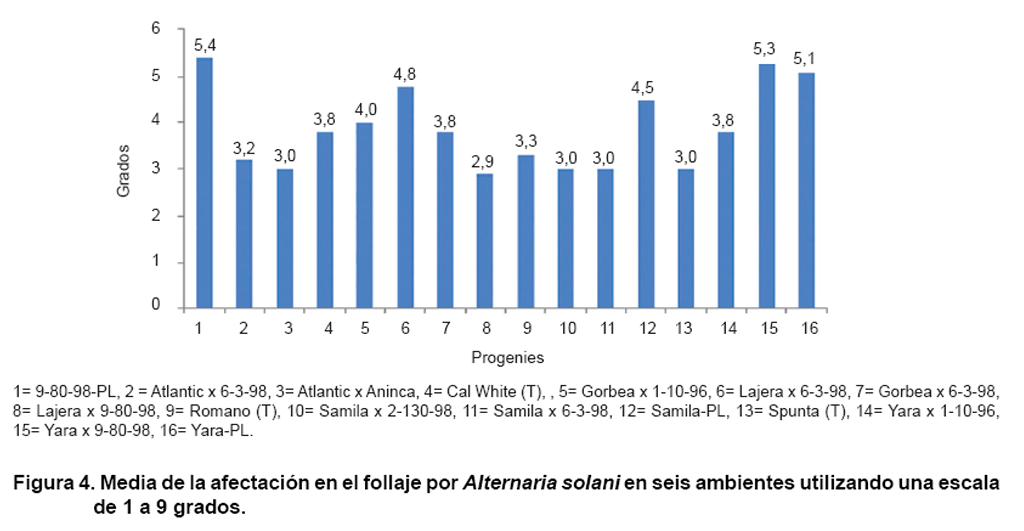
La Figura 4 ofrece una
visión general de la afectación por Alternaria solani
que causa la enfermedad del tizón temprano. Las progenies híbridas
8= Lajera x 9-80-98, 3= Atlantic x Aninca, 10= Samila x 2-130-9, 11= Samila
x 6-3-98 presentaron bajo grado de afectación, resultado que podría
estar indicando que los progenitores utilizados poseen genes de resistencia
y tolerancia, además de alta habilidad combinatoria para trasmitirla
a la descendencia.
En esta figura también se aprecia que las progenies de PL alcanzaron
los grados de afectación más elevados, resultados que pudieran
estar condicionados con la presencia de cierto grado de endogamia en este tipo
de material genético.
Este resultado podría estar influenciado por la disposición genética
de los cultivares que modifican en mayor o menor cuantía el ataque de
este hongo. La expresión de una resistencia también depende de
la presión de la enfermedad. Bajo condiciones de campo (natural) la presión
puede variar considerablemente de un año a otro y entonces algunas progenies
susceptibles pueden escapar a la infección y erróneamente aparentar
resistencia.
Este resultado también pudiera estar relacionado con las interacciones
hospedante-patógeno, las que son influenciadas por una interacción
genotipo-ambiente. La infección por A. solani que causa el tizón
temprano puede ser favorecido por estrés de sequía y calor, deficiencia
de azufre, manganeso o nitrógeno y senescencia temprana de la planta,
resultados que concuerdan con los encontrados cuando se analizó la resistencia
y la heredabilidad de la madurez de la planta en diferentes variedades de papa
(15, 16).
CONCLUSIONES
La progenie 6
(Lajera x 6-3-98) es promisoria para la producción de papa por su alto
y estable rendimiento y el rendimiento de tubérculos.
Las progenies 10 (Samila x 2-130-98), 14 (Yara x 1-10-96), 15 (Yara x 9-80-98),
3 (Atlantic x Aninca) y 11 (Samila x 6-3-98) se caracterizaron por su inestabilidad
en el número de tubérculos, en el rendimiento y en la masa promedio.
RECOMENDACIONES
Los contrastes en el comportamiento agronómico de las progenies de semilla sexual entre diferentes localidades se atribuyen a la interacción genotipo-ambiente, lo cual sugiere la necesidad de evaluar los materiales en cada zona de producción.
BIBLIOGRAFÍA
1. Organización
de las Naciones Unidas para la Agricultura y la Alimentación. Crops statistics
database. [en línea]. (2010). [Consultado: 5 octubre 2012]. Disponible
en: <http://faostat.fao.org/>.
2. Niño, L.; González, L.; Prieto, L.; Acevedo, E. y Suárez,
F. Producción de tubérculos semillas a partir de progenies de
semilla sexual de papa en Pueblo Llano, estado Mérida. Agronomía
Trop., 2010, vol. 60, no. 3, pp. 3-8. ISSN: 0002-192X.
3. Roa, S.; Barboza, C. y Zambrano, A. Estabilidad del rendimiento de variedades
de papa (Solanum tuberosum L.) para procesamiento industrial en el
estado Táchira, Venezuela. Rev. Fac. Agron. (LUZ). 2010. vol.
27, pp. 173-192. ISSN: 0378-7818.
4. Haynes, K. G.; Gergela, D. M.; Hutchinson, C. M.; Yencho, G. C.; Clough,
M. E.; Henninger, M. R.; Halseth, D. E.; Sandsted, E.; Porter, G. A. y Ocaya,
P. C. Early generation selection at multiple locations may identify potato parents
that produce more widely adapted progeny. Euphytica. 2012, vol. 186,
no. 2, pp. 573-583. ISSN: 0014-2336.
5. Haydar, A.; Islam, M.; Ara, T.; Khokan, E. y Hossain, M. Stability analysis
for tuber yield components in potato. Int. J. Sustain. Crop Prod. 2009,
vol. 4, no. 4, pp. 01-04. ISSN: 1991-3036.
6. Cuba-MINAG. Instructivo técnico para la producción de papa
en Cuba. Ed. Impresiones MINAG. La Habana. 2012. 59 pp.
7. Estévez,
A.; Salomón, J.; Castillo, J.; Ortíz, Ú. y Ortíz,
E. Regionalización de clones cubanos de papa (Solanum tuberosum
L.). Cultivos Tropicales. 2003, vol. 26, no. 3, pp. 59-63. ISSN: 1819-4087.
8. Varela, M.; Vicente, L.; Galindo, P.; Vázquez, A.; Castillo, J. y
Estévez, A. Una generalización de los modelos AMMI basada en el
algoritmo de TUCKALS3 para el análisis de componentes principales de
tres modos. Cultivos Tropicales. 2008. vol. 9, no. 1, pp. 69-72. ISSN:
1819-4087.
9. Sharma, A. K.; Venkatasalam, E. P. y Kumar, V. Potato mini-tuber production
during main and off crop seasons in high hills of north-western Himalaya. Potato
J., 2013. vol. 40, no. 1, pp. 29-37. ISSN: 1871-4528.
10. Navarro, J. A. Herencia y Ambiente. Boletín FEDAES. Barcelona. 2003.
22 pp. ISBN: 3-10-049512-8.
11. González, L.; Niño, L.; Villamizar, E.; Suárez, F.;
Acevedo, E. y Prieto, L. Evaluación de progenies de semilla botánica
de papa (Solanum tuberosum L.) en el Estado Mérida, Venezuela.
Bioagro, 2010. vol. 22, no. 3, pp. 12-21. ISSN: 1316-3361.
12. Pérez D. J.; González, A.; Sahagún, J.; Vázquez,
L. M.; Rivera, A.; Franco, O. y Domínguez, A. The identification of outstanding
potato cultivars using multivariate methods. Cien. Inv. Agr. 2009.
vol. 36, no. 3, pp. 391-400. ISSN: 0718-1620.
13. Yan, W.; Hunt, L. A.; Sheng, Q. y Szlavnics, Z. Cultivar evaluation and
mega- environment investigation based on the GGE biplot. Crop Science.
2000. vol. 40, pp. 597-605. ISSN: 0011-183X.
14 Argenpapa. Tizón Temprano-Síntomas y recomendaciones para su
control químico. Información Técnica. [en línea].
2013. [Consultado: febrero 2014]. Disponible en: <http://www.argenpapa.com.ar>.
15. Tiemens, H. M.; Delleman, J., Eising, J. y Van Bueren, E. Potato breeding,
un manual práctico para la cadena de la papa. Netherlands, 2013. 170
pp. ISBN: 978-90-802036-8-6.
16. Slater, A.; Wilson, G.; Cogan, N.; Forster, J. y Hayes, B. Improving the
analysis of low heritability complex traits for enhanced genetic gain in potato.
Theoretical and Applied Genetics. 2014, vol. 127, no. 4, pp. 809-820.
ISSN: 0040-5752.
Recibido: 1 de
marzo de 2014
Aceptado: 22 de diciembre de 2014
Ms.C. J. L. Salomón, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32700. Email: salomon@inca.edu.cu